Welke bewijzen zijn er voor evolutie vanuit de geologie?
Hoofdstuk 1 Inleiding.
Het is van belang om je te realiseren dat de wetenschap geologie een combinatie vormt van historische-en natuurwetenschap.
Historie binnen de geologie beschrijft situaties die korte tijd, of heel lang geleden, gebeurd zijn. Het is geen 'geschiedenis-wetenschap' omdat er geen geschreven bronnen beschikbaar zijn. Wetenschappelijke kennis van het verleden maakt slechts beperkte toepassing van 'bewijs door experiment' mogelijk.
Geologie leunt als natuurwetenschap sterk op de biologie omdat fossielen van vroeger levende organismen een leidende rol spelen. Daarnaast zijn natuurverschijnselen als klimaat, bodemgesteldheid, aardlagen, vulkanisme, aardbevingen, magnetisme, continentverschuivingen en zelfs kosmische evenementen van groot belang.
Het ontstaan van al die te bestuderen feitelijkheden is een heel apart hoofdstuk. De vraag naar ontstaan is een wezenlijk andere dan de vraag naar voortbestaan.
Toegepast op levende wezens betekent dit dat de vondst van fossielen niet kan bijdragen aan een theorie over het ontstaan van die organismen. Je kunt wel onderzoeken hoe een gestorven organisme veranderen kan op een specifieke plek in een aardafzetting tijdens een bepaalde periode maar dat levert geen bijdrage aan een theorie over de evolutie van die soort of dat geslacht vanuit een vroeger levend organisme.
De biologie is hulpwetenschap voor de geologie!
Alleen de biologie kan het ontstaan van soorten levende wezens proberen te verklaren (als ze daar de middelen en methoden voor beschikbaar heeft!).
Bij de vraag naar de mogelijke evolutie van de mens hebben we al diverse geologische "bewijzen" ter sprake gebracht (zie pg. evolutie II).
Ik wil nu proberen tot een heel voorlopig overzicht te komen wat er in de wetenschap van de geologie bekend is over evolutie van organismen. Omdat ik geen geologie gestudeerd heb vergt dit voor mij nieuwe kennis ontwikkelen.
Mijn eerste kennismaking met imponerende levensgrote voorbeelden van Dinosauriërs betroffen de Iguanodonten van Bernissart.
Ze waren tentoongesteld in het Natuurhistorisch Museum te Brussel. Als ik me goed herinner betrof het een verzameling van ong. 30 geweldige dino's die samengeperst in een diepe laag zandsteen van een kolenmijn waren begraven. Dit beeld van meerdere grote dinosauriërs, ontdekt in een betrekkelijk klein gebied, komt herhaaldelijk voor. Ik hoop daar verderop v.b. van te geven.
De eerste vraag die daarbij steeds weer opduikt is: hoe kunnen de skeletten van deze dieren, die volgens de evolutieleer ong. 65 miljoen jaar en langer geleden op aarde rondzwierven, zo bij elkaar geworpen, bewaard zijn gebleven. De plaatsing in een kolenmijn wijst er m.i. op dat de vondsten ontstaan zijn door orogeneseprocessen.
Deze vraag is dezelfde bij de duizenden skeletten van de langharige mammoeten die geconserveerd zijn in de Siberische permafrost. Nog altijd trekken bewoners van die gebieden er in hun voorjaarsvakanties op uit om slachttanden van goede kwaliteit op te sporen en voor enkele duizenden dollars per stuk te verhandelen.
In de loop van enkele eeuwen zijn er duizenden tonnen aan ivoor van mammoeten voor veel geld in omloop gebracht.
Overbekend is dat er vele mammoeten zijn gevonden met nog "verse" voedingsresten in hun magen en tussen de tanden. Hoe kunnen dieren van deze grootte in zo korte tijd zijn diepgevroren?
Diverse oorzaken zijn daartoe bedacht maar geen daarvan overtuigt. Voor de 'Iguanodonts de Bernissart' geldt m.i. hetzelfde. Werden ze meegesleurd door een grote vloedgolf en meteen daarna bedekt door meters zand/klei? Of speelde een aardbeving en/of vulkaanuitbarsting een rol?
Al met al vermoed ik dat bestaande theorieën over vindplaatsen en vorming van fossielen opnieuw getoetst zouden moeten worden op deugdelijkheid. Op grond van bewijsbare feiten, aangevuld met experimenten.
Als het gaat over de vondsten van fossielen in kolenlagen, ontstaan in het Carboon, Perm en zelfs Trias zien we overeenkomstige problemen die om beantwoording vragen. Hoe en waaruit kunnen honderden meters dikke steenkoollagen worden gevormd in vrij korte tijd? Dit laatste is noodzakelijk omdat anders de organische plantenmaterialen waaruit steenkool is gevormd volledig worden opgeruimd door schimmels en bacteriën. Zeker in het tijdperk van het Carboon toen over het grootste deel van de aardbol een tropisch klimaat aanwezig was.
Om het nog moeilijker verklaarbaar te maken: diverse boomstammen doorbreken vele kolenlagen. En staan daarbij vaak op hun kop; de kruinen naar beneden, de wortels naar boven.
In ieder geval is het grote adagium van de geologie, ontwikkeld door Lyell, niet te handhaven.
Lyell meende ontdekt te hebben dat we voor de verklaring van geologische processen dienen uit te gaan van de huidige processen die in de natuur plaats vinden. Het zgn. actualietisprincipe, ook wel genoemd het uniformitarianisme.
Dus verwering, sedimentatie, gebergtevorming en continentverschuiving vinden zeer geleidelijk plaats. Deze 'leer' werd verdedigd tgo. Cuvier die van mening was dat grote catastrofes een overheersende rol hadden gespeeld tijdens de vorming van de aardlagen. Cuvier ging zelfs uit van elkaar opvolgende scheppingsperioden.
Intussen is het principe van Lyell wel behoorlijk ondermijnd door geologen. Alleen al de veronderstelling dat het tijdperk van de dino's werd veroorzaakt door een ramp met gevolgen voor het hele leven rond de aardbol maakt dit duidelijk. De inslag van een kolossale meteoriet in de omgeving van Mexico wordt door velen gezien als de oorzaak van het verdwijnen van de grote dino's op aarde.
Een inslag van zo grote omvang dat het hele klimaat werd verstoord en het leven op aarde ter nauwer- nood werd gered.
Hoofdstuk 2
De evolutie van de belangrijkste groepen organismen.
2.1 Bacteria, archaea en eukarya.
Gegevens over bouw, functioneren en indeling ontleen ik vooral aan het naslagwerk:
Microbiology, fifth edition door Prescott, Harley en Klein.
uitg. McGraw-Hill. 2002.
Bacteriën komen overal voor. Voedingsbron en milieuomstandigheden (temp., licht, zuurgraad) bepalen welke soort waar voorkomt. Onze huid zit vol bacteriën. In ons darmkanaal is de zgn. bacterieflora aanwezig: miljarden bacteriën in een al of niet evenwichtige samenstelling. Een verstoorde darmflora kan allerlei gezondheidsproblemen veroorzaken.
Fossiele microben zijn vooral te vinden in Stromatolieten. Dikke stukken Stromatolieten vind je aan de kusten van West-Australië. De lagen zijn en worden gevormd door cyanobacteriën (blauwalgen) en andere micro-organismen. Deze organismen incorporeren calciumsulfaten en calciumcarbonaten.
Deze fossielen worden gedateerd op 3.5 miljard jaar geleden.
Andere soorten bacteriën worden geschat op 1.5 miljard jaar (gevonden in het Oeralgebergte) of op 950 miljoen jaar (gevonden in Oost-Siberië).
Door het ontbreken van grote hoeveelheden fossielen is het onmogelijk om een ontwikkelingslijn van bacteriën en archaea op te stellen.
Men veronderstelt dat de eerste prokaryoten onder anaerobe omstandigheden, dus zonder zuurstof, leefden. De cyanobacteriën of blauwalgen worden wel beschouwd als oorsprong van de prokaryoten.
Men onderscheid vaak drie grote domeinen in de levende natuur: bacteria, archaea en eukaria.
De archaea hebben bijzondere vetten in hun celmembraan (lipiden van diglycerol-tetra-ether of isopreenglycerolen-di-ether) en bijzonder rRNA (ribosoom-RNA) in de cel.
De bacteria bezitten celmembranen met lipiden van diacyl-glycerol-diesters en bacterieel rRNA.
De eukaria hebben membranen met lipiden van glycerol-acyldiëster en eucaryotisch rRNA.
Tot de eukaria (eukaryoten) behoren de schimmels, algen, planten, dieren en mensen (volgens de oudere indeling).
Er bestaan diverse speculaties omtrent het ontstaan van deze drie domeinen. De celmembranen van de eerste levende cellen kunnen ingevouwen zijn en dubbele membranen hebben gevormd, waaruit langzamerhand mitochondriën, chloroplasten en andere organellen zijn ontwikkeld. Deze eerste, primitieve organellen moeten dan wel stukjes DNA bij zich gehad hebben.
Een andere onbewezen hypothese is dat een fusie tussen bacteria en archaea een eerste begin van een eucaryotische cel vormde. Dan moet er nog wel een celkern ontstaan zijn. Daarbij natuurlijk ook het verlies van de typische bacterie-celwand en de vorming van endoplasmatisch reticulum. Intussen natuurlijk nog de oude genen kwijt raken en nieuwe vormen!
Mitochondriën en chloroplasten kunnen later gevormd zijn door veronderstelde symbiose met fotosynthese-bacteriën.
Prochloron schijnt hiervoor een goede kandidaat te zijn. Dit organisme leeft als extracellulaire symbiont op koraalriffen en bevat chlorofyl a en b. Het kan CO2 als bouwstof incorporeren via de Calvincyclus van de fotosynthese. Net als groene planten doen. Deze kandidaat wordt echter door vele wetenschappers afgewezen als stap op weg naar groene algen. Omdat er geen fylogenetische tussenstappen zijn.
Alle mogelijke vormen van stofwisseling komen bij bacteria en archaea voor:
- aeroob of anaeroob
- autotroof of heterotroof
- anaerobe fotosynthese
- chemo-organotroof.
Er zijn meer dan 6 verschillende "stambomen" over ontwikkeling en vervlechting van de drie domeinen bedacht. Chloroplasten en mitochondriën duiken daarin op dankzij endosymbiose met cyanobacteriën. Door dit "wonder" ontstaan in planten, dieren en schimmels de mitochondriën en/of chloroplasten. Voor deze hypothese bestaat geen onderbouwing vanuit fossielmateriaal!
Conclusie: de natuurwetenschap heeft geen idee waar eukaryote cellen vandaan komen!
(evenmin waar bacteria en archaea vandaan komen).
Cyanobacteriën (blauwalgen) worden vaak genoemd als bron van de evolutie naar andere protozoa en groene planten. Cyanobacteriën zijn wel heel wonderlijk gebouwd. Ze bevatten plastiden die CO2 kunnen binden en omzetten in O2. Het chlorofyl in cyanobacteriën is eenvoudiger gebouwd dan het chlorofyl in groene planten. Om het zonlicht door te geven voor de fotosynthese zijn de pigmenten fycoerytrine en fycocyanine noodzakelijk aanwezig. Deze pigmenten zijn weer een onderdeel van het ficobilisoom. Dit laatste bestaat uit eiwitcomplexen. Het geheel vergt dus een lange reeks van assimilatiestappen vanaf het DNA, via RNA's naar mitochondriën. Zoals intussen welbekend!
Blauwalgen zijn berucht omdat ze warm en voedselrijk zwemwater kunnen verzieken. Ze veroorzaken dan irritaties aan ogen en huid. Ook veroorzaken ze hoofdpijn en maag- en darmklachten.
2.2 Protozoa.
Protozoa zijn 1-cellige organismen. Het zijn eukaria zonder chloroplasten. Ze zijn dus heterotroof. Meestal hebben ze 1 of meerdere zweepharen of trilharen voor de voortbeweging. Ze leven van andere 1-celligen of bacteriën. Of ze leven parasitair in vele soorten organismen. Protozoa kunnen een zeer ingewikkelde cyclus van geslachtelijke voortplanting vertonen. Eerst vindt er conjugatie van b.v. twee pantoffeldiertjes plaats (1). Daarbij verdwijnen eerst macronuclei gedeeltelijk en delen de micronuclei zich door meiose in vieren (3). Drie van de vier verdwijnen weer. Daarna deelt de overgebleven haploïde nucleus zich (5). Daarop vindt er uitwisseling plaats twee haploïde kernen (6). De beide diertjes scheiden zich en de ontstane diploïde nucleus in beide diertjes deelt zich drie keer door mitose (10, 11). Drie van de acht diploïde nuclei verdwijnen en vier van de acht vormen samen een nieuwe macronucleus (13).
Onmogelijk om hieruit een evolutie-verhaal tevoorschijn te toveren dat duidelijk kan maken hoe geslachtelijke voortplanting annex "liefdesuitingen" kan zijn ontstaan.
2.2.1 De Archezoa zijn 1-cellige eukaryotische organismen maar zonder Golgiapparaat, mitochondriën en chloroplasten. Voorbeeld Giardia. Het is een parasiet die leeft in de darmen van zoogdieren en mensen.
https://upload.wikimedia.org/wikipedia/commons/thumb/0/07/Giardia_life_cycle_nl.svg/400px-Giardia_life_cycle_nl.svg.png
2.2.2 Diatomeeën zijn 1-cellige algen (dus ook eukaryoten) die opgesloten zijn tussen 'doosje-dekseltje'.
Vgl. twee petri-schaaltjes. Na elke celdeling wordt het kleinste deksel steeds kleiner. Door een tussenstadium van geslachtelijke voortplanting in te voegen groeit de zygote weer uit tot de oorspronkelijke grootte.
https://th.bing.com/th/id/OIP.aNtabAu4zrCdgd5O_blfIgHaKd?w=116&h=180&c=7&o=5&pid=1.7
Diatomeeën vormden dikke lagen diep op de zeebodem of in meren. Het bestaat uit kiezelslik (SiO2) en wordt per zak verhandeld. Het is voor vele nuttige doeleinden bruikbaar.
Ze zijn een rijke voedingsbron voor zoöplankton, kreeftjes, schelpdieren, vissen enz.
In Nederland werd het tijdens het holoceen gevormd. Dus vrij kort geleden (11.700jr.).
Volgens Lehrbuch der Botanik (Stuttgart 1967) dateren de oudste vondsten uit de Jura, ong. 200 miljoen jr. geleden. Vrij laat dus in de aardgeschiedenis. Oftewel: vanaf de schepping. Zie daarvoor de pg. Bijbels georiënteerde geologie.
Verlaten groeve voor diatomeeënaarde.
https://upload.wikimedia.org/wikipedia/commons/thumb/a/a0/KieselgurOhe.jpg/200px-KieselgurOhe.jpg
2.2.3 De enige protozoa die een belangrijke rol spelen in afzettingen van aardlagen zijn, voorzover ik weet, de Foraminiferen en de Radiolariën. Ze hebben vaak heel diverse en prachtig gebouwde skeletjes, vergelijkbaar met diatomeeën.
Foraminiferen

.
https://upload.wikimedia.org/wikipedia/commons/thumb/b/b8/Ammonia_beccarii.jpg/266px-Ammonia_beccarii.jpg
Foraminiferen zijn belangrijke gidsfossielen. Hun kalkskeletjes bestaan uit verschillende kamertjes. Ze komen al voor in Cambrische aardlagen. Drie à vier miljard jaar (?) oud.
Radiolariën
https://upload.wikimedia.org/wikipedia/commons/thumb/6/6a/Haeckel_Acanthophracta.jpg/266px-Haeckel_Acanthophracta.jpg
Radiolariën hebben kiezelskeletten, net als diatomeeën. Radiolariën komen ook in Cambrische gesteenten voor. Ze vormen soms lagen van enkele tientallen of zelfs honderden meters dik.
Vergelijk maar met de kalkrotsen van Zuid-Engeland. Hoeveel tijd zou er nodig zijn om zulke pakketten te vormen?
2.3 Fungi (schimmels).
Schimmels zijn heterotroof omdat ze geen bladgroenkorrels bezitten. Ze produceren daartoe exo-enzymen die de organische stoffen buiten de cellen verteren. Daarna absorberen ze de oplosbaar gemaakte stoffen. Ze planten zich geslachtelijk of ongeslachtelijk voort. De verspreiding vindt meestal door sporen plaats.
Vele leden van de afdeling Basidiomyceten vormen bovengronds de welbekende paddenstoelen.
De ondergrondse schimmeldraden die uit een ontkiemende spore groeien zijn geslachtelijk verschillend en haploïd. Ze zijn uiterlijk niet van elkaar te onderscheiden. Ze worden aangegeven met + of - draden. Als + en - elkaar tegen komen gaan ze fuseren maar de kernen versmelten niet met elkaar. Er vormt zich dus geen zygote. Nadat door een wonderlijk groeiproces een paddenstoel gegroeid is vormen zich op de plaatjes de basidiosporen. Binnen deze sporen vindt samensmelting van beide kernen plaats. De dan ontstane zygote gaat zich delen door meiose en vormt daarbij 4 haploïde celkernen. De haploïde sporen (miljarden, biljoenen, biljarden of triljarden) verspreiden zich en vormen weer de + en - draden.
https://upload.wikimedia.org/wikipedia/commons/thumb/b/bc/03_02_09_life_cycle_of_Agaricus_sp.%2C_Agaricales_Basidiomycota_%28M._Piepenbring%29_nl_txt.png/250px-03_02_09_life_cycle_of_Agaricus_sp.%2C_Agaricales_Basidiomycota_%28M._Piepenbring%29_nl_txt.png
Schimmels zijn vaak erg belangrijk voor het leven van andere organismen: driekwart van alle vaatplanten leven samen met schimmels (mycorrhiza) die een dicht netwerk vormen rond hun wortels. De schimmel ontvangt organische voedingsstoffen van de groene plant en levert de nodige anorganische stoffen voor de bomen en andere planten. Orchideeën kunnen helemaal niet ontkiemen of groeien zonder hun mycorrhiza.
Fungi zijn essentiële onderdelen voor fermentatieprocessen bij de bereiding van brood, wijn en bier e.a. levensmiddelen. De eencellige gisten spelen hierin de hoofdrol. Gisten planten zich snel voort door knopvorming en produceren daarbij veel CO2.
Van enorm belang is de productie van antibiotica (vooral penicilline) door schimmels.
De celwand van schimmels bestaat o.a.uit chitine, dezelfde stof als het skelet van insecten.
De oudste schimmels zijn ong. 1.4 miljard jr. geleden ontstaan, menen de geleerden.
Tijdens het Cambrium (ong. 500 miljoen jr.) veroverden ze het land.
Tijdens het Devoon (ong. 400 miljoen jr. geleden) waren ze heel algemeen.
Basidiomyceten en Ascomyceten (met de zakvormige ascus als voortplantingsorgaan) ontstonden in die tijd uit een gemeenschappelijke voorouder. Die voorouder moet nog gevonden worden.
Vanaf het Carboon (350 miljoen jr. geleden) zijn alle klassen schimmels aanwezig.
Over het ontstaan van de verschillende stammen en klassen is weinig of niets met enige zekerheid bekend.
2.4 Algae.
Algen zijn 1-cellige planten of ze zijn bladvormig. Meestal zijn het lange draden met bladgroenkorrels. Eencellige algen vind je veel op stenen, hout en boomstammen. Aan het strand vind je bladvormige wieren (zeesla). Vaak zie je daar ook bruin- en roodwieren. In de Grote Oceaan, aan de westkust van Noord-Amerika, groeien enorme wieren (kelp) die meebewegen met de stromingen in het water. Vele zeedieren vinden daartussen een prima woonplaats.
Tot de Algae worden gerekend:
- de chlorophyta (de groene algen)
- de euglenophyta (met euglena, het groene oogdiertje)
- de dinoflagellaten (1-celligen met 2 zweepharen)
- de bruinwieren (de phaeophyta)
- de roodwieren (de rhodophyta)
- de diatomeeën .
- Volvox (een bolvormige kolonie van cellen).
Met als veronderstelling dat alle bacteriën, protozoën en algen, zijn ontstaan uit de universele voorouder! Die nooit zal worden gevonden??
In de pg. 'Een Bijbels georiënteerde geologie' zal ik proberen duidelijk te maken dat deze groepen organismen vanaf het begin van de schepping aanwezig waren in de landschappen van het Pangea.
Samen met alle hoofdgroepen van het plantenrijk en het dierenrijk.
Korstmossen (beter: korstmorsen) bestaan uit een hechte vervlechting van schimmel en alg.
Je vindt ze overal op stenen en bomen.
a = bovencortex b = algenlaag c = merg d = ondercortex e = aanhangsels.
https://upload.wikimedia.org/wikipedia/commons/thumb/e/ed/File-Meyers_b6_s0351a.jpg/260px-File-Meyers_b6_s0351a.jpg
De groene alg produceert dankzij fotosynthese de organische stoffen.
De schimmel beschermt, houdt water vast en levert de anorganische voedingsstoffen die ze opneemt dankzij de uitscheiding van zuren.
Korstmorsen groeien uitermate langzaam. Ze kunnen worden gebruikt om de ouderdom van gesteenten te bepalen.
LATER ZULLEN WE ZIEN DAT DE NIEUWE INDELING VAN DE ORGANISMEN OP BASIS VAN DE CLADISTIEK EEN GEHEEL ANDERE GROEPERINGSVORMEN TOT GEVOLG HEEFT.
2.5 Mossen varens en paardenstaarten.
Mossen, paardenstaarten en varens behoren tot de meest bekende sporenplanten.
Ze zijn echter totaal verschillend gebouwd.
2.5.1 Mossen hebben geen vaatbundels. Ze hebben daarom geen echte wortels, stengels en bladeren.
Mosblaadjes zijn eenvoudige parenchymweefsels met veel bladgroenkorrels zonder epidermis of cuticula. Een mosblaadje kan een "nerf" hebben die 2 of 3 cellagen dik is maar geen vaten bevat.
Ze drogen daarom snel uit maar leven heel snel weer op als er vocht bij komt. De "worteltjes" zijn draadvormig (rhizoiden). Ze bestaan uit 1 cellaag.
https://upload.wikimedia.org/wikipedia/commons/thumb/9/92/Moss_life_Cycle.jpg/350px-Moss_life_Cycle.jpg
De afbeeldingen hierboven laten zien hoe een mosplantje groeit en zich voortplant.
Uit een haploïde spore groeit een kiemdraad waaruit al gauw een mosplantje ontstaat.
In de top van het plantjes ontstaan mannelijke (antheridium) en vrouwelijke (archegonium) voortplantingsorgaantjes. Als er maar een waterdruppel aanwezig is zwemmen de spermatozoïden naar de eicellen. Uit de zygote groeit de sporofoor. Deze blijft op het plantje staan en vormt een steeltje ( de seta) met daarop een sporekapsel (de capsule). In het sporekapsel ontstaan na meiose de haploïde sporen. Na ontkieming van de spore is de cirkel weer rond.
Samengevat: het mosplantje is de gametofyt. Alle cellen zijn haploïd.
Het sporenkapsel ontstaat na bevruchting. Alle cellen zijn diploïd.
Levermossen (Hepaticae) groeien op veel plaatsen tussen stenen en tegels. Het zijn bladvormige structuren die voortplantingsorgaantjes vormen op het "blad".
Levermossen met broedbekertjes.
Ze worden beschouwd als de eerste landplanten. Ontstaan ong. 475 miljoen jr. geleden ? In het Ordovicium.
Van mossen wordt gezegd dat ze in het Devoon zijn ontstaan (400 miljoen jr. geleden).
Mossen en levermossen leefden waarschijnlijk samen op Pangea.
Even je realiseren wat er allemaal nodig is om een mosplantje te doen functioneren in vergelijking met een schimmel of alg. Alleen de eerste stap, de groei van een draadje uit de spore lijkt op vorming van een alg of een schimmel.
Vervolgens zijn er honderden of duizenden genen nodig om de groei van gametofyt en sporofyt in goede volgorde te realiseren. Hoe worden gewone celdelingen, van haploïde of diploïde cellen, nauwkeurig afgewisseld met reductiedelingen en bevruchting door gameten?
Evolutiebiologen waren en zijn gefocussed op genen en eiwitten. Veel DNA werd daarom beschouwd als junk-DNA, een overbodig restant van het evolutieproces. DNA, samen met de drie soorten RNA's, zijn echter noodzakelijk voor de volgorde in de hele cyclus van groei en ontwikkeling. Bovendien moeten ze zorgen voor de opbouw van vele andere organische stoffen (cellulose, glycogeen etc. etc.) En dat alles in de goede volgorde. en in juiste hoeveelheden.
2.5.2 Varens behoren tot de vaatplanten. Ze hebben vaak grote, echte bladeren met nerven waarin zich hout- en bastvaten bevinden. Hout- en bastvaten groeien vanuit het cambium. Zo ontstaat de diktegroei bij alle vaatplanten. Varens hebben hun sporangia, vaak in groepjes bij elkaar op de bladeren liggen.
Varens hebben ook echte wortels. En vaak (ondergrondse) wortelstokken.
 Sporangia aan de onderzijde van een varenblad. Ze zijn vaak bedekt met een vlies om de sporangia te beschermen. De sporangia hebben een annulus om ze open te scheuren bij droogte om de sporen vrij te laten.
Sporangia aan de onderzijde van een varenblad. Ze zijn vaak bedekt met een vlies om de sporangia te beschermen. De sporangia hebben een annulus om ze open te scheuren bij droogte om de sporen vrij te laten.
De generatiewisseling is bij varens heel anders dan bij mossen. De varenplant is de diploïde sporofyt en de gametofyt is een klein, kort levend, bladachtig groen orgaantje dat op de grond ligt en de gameten vorm. Na de bevruchting groeit uit de voorkiem de jonge varenplant.
https://upload.wikimedia.org/wikipedia/commons/4/44/Varen.png
Er bestaan ook boomvarens maar dat zijn varens met een schijnstam omdat ze wel een boomachtig uiterlijk hebben maar ze bestaan uit samengroeiende bladstelen.
Fossiele varens worden gevonden in Devoon en Carboon. An het einde van het Perm sterven de meeste varens uit. Ze komen massaal terug vanaf Jura en Krijt. Dan verschijnen ook de zaadplanten (angiospermen). Het zijn tegelijkertijd de perioden van de Dinosauriërs.
Aan het einde van het Perm vindt er massale uitsterving (extinctie) van bijna alle toen levende organismen plaats. Wel 90%. Dit herhaalt zich aan het einde van het Krijt.
Het zal duidelijk zijn dat er van een bewijsbaar evolutieproces van varens vanuit mossen geen sprake kan zijn.
Een vaatbundel, zoals bij alle vaatplanten voorkomt, is een gigantisch ingewikkeld bouwwerk dat door alle onderdelen van de plant loopt vanaf de wortelpuntjes tot in de uiteinden van de bladeren.
De houtvaten (xyleem) vervoeren water met daarin opgeloste anorganische stoffen naar boven. Het zijn dode cellen. Ze ontstaan uit levende cellen waarvan cytoplasma, celkernen en de cellulosewand verdwijnen. In de wanden ontstaat lignine.
De zeefvaten (floëem) zijn langwerpige , levende cellen die met elkaar verbonden zijn door zeefplaten.
Zeefplaten hebben een normaal gebouwde begeleidende cel. Dankzij de zeefplaten, met vele kleine openingen, kunnen organische stoffen naar boven of beneden worden getransporteerd; voor de groei van stengels, bladeren, bloemen, zaden, vruchten en wortels.
Bastvaten en zeefvaten ontstaan vanuit het cambium. Cambiumcellen delen zich gedurende een bepaalde periode bij de meeste soorten planten of jaren achtereen bij bomen.
Ligging, structuur en vorm van vaatbundels in de vaatplanten verschillen op duizenden manieren.
Stel je nu eens voor hoe vaatbundels zouden kunnen zijn ontstaan. In een "mosplantje" ontstaan enkele houtvaatjes. Daarvoor moet heel wat gebeuren (zie boven). De daarvoor nodige triplets in het DNA en RNA's moeten doorgegeven worden aan de volgende generatie. De houtvaten moeten verder evolueren en door het hele plantjes groeien. Daarvoor moeten de rhizoiden omgebouwd worden. Daarna de vorming van zeefvaten samen met hun begeleidende cellen, netjes naast de houtvaatjes. Maar dit kan alleen een samenhangend geheel worden als er tegelijkertijd cambiumcellen ontstaan. In een perfecte ligging tussen de hout- en zeefvaten en door alle onderdelen van het plantje heen. Start en beëindiging van de groei moeten nauwkeurig geregeld worden. Als al deze processen plaats gevonden hebben is er nog lang geen varenplant ontstaan. Daarvoor moet de hele voortplantingscyclus worden omgevormd!
Tegelijkertijd moeten er sporenkapsels op de nieuw gevormde bladeren worden aangelegd. Vaak moeten er rhizomen (wortelstokken) worden gebouwd zodat er door ongeslachtelijke voortplanting nieuwe varens kunnen groeien.
Geen bioloog kan dit grootse verhaal verklaren vanuit een gedetailleerde kennis van alle macromoleculen die nodig zijn en genetische processen die hiervoor vereist zijn. Bovendien is doelgerichte, teleologische ontwikkeling binnen de natuurwetenschappen verboden gebied. Natuurfilosofen en daarna theologen weten hier wel raad mee. Al fantaserend ontstaan er gemakkelijk echte stengels, bladeren en wortels uit primitieve (een kernbegrip in de evolutiebiologie) voorstadia (die niemand kent). Als het echt te moeilijk wordt kunnen theologen hun 'godsbegrip' toepassen op de niet te bevatten gebeurtenissen.
2.5.3 Paardenstaarten (Equisetae).
Overbekend, als lastig onkruid in bijna elke tuin, is heermoes.
Deze sporenplanten hebben geen bladeren, wel schubben die een kokertje rondom de stengel vormen.
De sporangiën bevinden zich aan de toppen van de stengels. Ze groeien vaak onder een schildje dat een zeshoekig afdakje vormt.
Net als bij varens ontwikkelen zich hierin sporen door reductiedeling (meiose). De sporen hebben 2 elateren die een luchtreis mogelijk maken. Als ze zich weer oprollen tijdens de reis valt de spore op een hopelijk vochtige plek. Goed geregeld dus. Na ontkieming ontstaat er een prothallium net als bij een varen. Na de bevruchting ontstaat er de zygote. De paardenstaart die hieruit ontstaat is dus ook diploïd.
Equisetum arvense (akkerpaardenstaart of heermoes) heeft in het voorjaar aparte stengels met sporangiën.

https://upload.wikimedia.org/wikipedia/commons/thumb/9/9e/Equisetumarvense.jpg/266px-Equisetumarvense.jpg
Heermoes, na de fertiele stengels, groeien de groene stengels met bladgroen en stengelomvattende,
schubvormige blaadjes.
2.6 Zaadplanten (naaktzadigen en bedektzadigen;
gymnospermen en angiospermen).
Alle zaadplanten produceren zaden die door een zaadhuid helemaal omgeven zijn. Dit betekent dat een zaadcel niet zomaar bij een eicel kan komen om daarmee samen te smelten. Daarvoor is nodig dat de stuifmeelkorrel eerst ontkiemt en een stuifmeelbuis vormt. In de stuifmeelbuis wordt de zaadkern gevormd die naar de microscopisch kleine opening in de zaadhuid moet worden gebracht.
2.6.1 Bij de Gymnospermen liggen de zaden op een vruchtschub. De vruchtschubben samen vormen een kegel (meestal verhoutend zoals bij dennen en sparren), of een bes (Jeneverbes). Bij Taxus is het zaad gedeeltelijk omgeven door de fel rood gekleurde arillus.
Als de kegels door droogte open springen kunnen de vaak gevleugelde zaden door de wind worden verspreid. Bij de Taxus zorgen vogels voor de verspreiding. De blauwachtig gekleurde jeneverbessen worden door lijsters verspreid.
Den met mannelijke en vrouwelijke kegels, vruchtschubben, zaden en kiemplant.
https://upload.wikimedia.org/wikipedia/commons/thumb/0/0b/Pinus_sylvestris_-_K%C3%B6hler%E2%80%93s_Medizinal-Pflanzen-106.jpg/266px-Pinus_sylvestris_-_K%C3%B6hler%E2%80%93s_Medizinal-Pflanzen-106.jpg
Jeneverbes.
https://upload.wikimedia.org/wikipedia/commons/thumb/f/fb/Jeneverbes_bessen.jpg/266px-Jeneverbes_bessen.jpg
Taxus met jonge en rijpe vruchtjes.
Het zaad met niet giftige arillus.
De bladeren bij de naaktzadigen bestaan meestal uit naalden. Bij sommige geslachten zijn het schubben, b.v. bij Cupressus, Thuja en Chamaecyparis.
2.6.2 Bij de Angiospermae, de bedektzadigen, zit de zaadknop opgesloten in het vruchtbeginsel. Het vruchtbeginsel is onderdeel van de stamper. De stamper bestaat uit stempel, stijl en vruchtbeginsel.
De stuifmeelkorrels moeten ontkiemen op de stempel van dezelfde soort bloem. Uit de stuifmeelkorrel groeit de stuifmeelbuis die door de stijl moet groeien om bij de zaadknop te arriveren. Tijdens die groei ontstaan hierin twee zaadkernen.
In de zaadknop bevinden zich meerdere cellen en kernen. Eén daarvan is de eikern. Twee andere kernen worden ook bevrucht nadat ze eerst zijn samengesmolten.
Bij de Angiospermae vindt dubbele bevruchting plaats:
1. één zaadkern smelt samen met de eikern. Hieruit ontstaat de diploïde
zygote. Uit de laatste ontwikkelt zich het embryo, de kiem. De kiem
blijft opgesloten in het zaad dat zich ontwikkelt uit de zaadknop.
2. een tweede kern smelt samen met de diploïde kern die al aanwezig is
in de zaadknop. Uit de aldus gevormde triploïde kern groeit het
reservevoedsel dat nodig is voor de groei van de kiem tijdens de
ontkieming.
Bij de tweezaadlobbigen bevindt de kiem zich tussen 2 zaadlobben. Als je pinda's, bruine bonen of doperwten eet kun je dat allemaal goed zien. Tweezaadlobbige planten hebben veernervige of handnervige bladeren.
Bij de éénzaadlobbigen ligt de kiem aan het uiteinde van de zaadlob. Eenzaadlobbige hebben lijnnervige of paralelnervige bladeren. Denk maar aan grassen, zegges, orchideeën enz.
De voortplantingsorganen (stampers en meeldraden) bij de bedektzadigen zijn meestal omgeven door een perianth. Dit perianth bestaat meestal uit kelk- en kroonbladeren. Eén van de twee of beide kunnen afwezig zijn of sterk gereduceerd. Vooral bij windbloeiers is dit laatste vaak het geval omdat het lokken van insecten niet nodig is. Windbloeiers maken altijd grote hoeveelheden en licht stuifmeel. Denk maar aan bloeiende grassen, elzen, berken etc.
2.7 Bouw en voortplanting van bovengenoemde groepen.
Als we nu dit hele hier boven vertelde verhaal proberen te overzien kan duidelijk zijn dat er ontwikkelingslijnen voorstelbaar zijn als het gaat om de bouw van de planten met stengels, wortels en bladeren. Eveneens zijn er ontwikkelingslijnen denkbaar in de wijze van geslachtelijke voortplanting.
Maar voorstelbaarheid in de fantasie is wezenlijk iets anders dan bewijzen in de (natuur)wetenschap!
Ongeslachtelijke voortplanting speelt bij alle plantaardige organismen een grote rol. Gewoon door celdeling of m.b.v. speciale organen zoals wortelstokken, uitlopers, knollen, bollen enz.
Als we letten op de vegetatieve bouw valt het volgende op: algen en schimmels hebben geen vaatbundels, bladeren enz. Ze leven zowel op het land als in het water. Ze zijn wel principieel verschillend gebouwd. Denk aan de tweekernige schimmeldraden, de vorming van paddenstoelen of de asci van de Ascomyceten. Er is geen enkel steekhoudend bewijs voor de evolutie van beide afdelingen van deze organismen.
De Bruinwieren kunnen erg groot worden (tot wel 100 m. lang) met bijna boomvormig uiterlijk. Hun uitgroeisels doen dan denken aan wortels, stengels en bladeren. Ze hebben bruine chromatoforen (fucoxanthine).
Dan is er nog de zeer bijzondere afdeling Lichenen of Korstmorsen. Altijd landplanten op plaatsen onder extreme omstandigheden: op muren, boomstammen, rotsen ........ Er bestaan meer dan 20.000 soorten Lichenen.
Het ontstaan van de complexe bouw van schimmel en alg samen is nooit door fossielen geboekstaafd.
Terwijl ze toch op zoveel plaatsen over bijna de hele aardbol voorkomen!
Bij de volgende Afdelingen is er een volgorde in bouw op te merken.
De mossen leven zonder hout- en bastvaten, huidmondjes of cuticula. Toch echte landplantjes.
Ze zouden ontstaan kunnen zijn in het Ordovicium of Devoon.
Maar: "gebrek aan fossielen maakt gedachten omtrent hun fylogenese nog speculatiever dan die van hogere planten". Aldus in Strasburger; Lehrbuch der Botanik.
De varens hebben echte houtvaten en bastvaten maar nog geen secundaire diktegroei. Hun vaak grote bladeren bezitten een uitgebreide nervatuur. Vaak hebben ze wortelstokken.
Ze kunnen ontstaan zijn in Devoon en zijn sterk aanwezig in Carboon.
De paardenstaarten zijn totaal anders gebouwd. De stengels zijn geleed en hebben op de tussenschijven een krans van schubvormige, kleine, vergroeide blaadjes. Er zijn wel vaatbundeltjes. En vaak wortelstokken. Bij uitgestorven boomvormige paardenstaarten kwam secundaire diktegroei voor.
Paardenstaarten zijn al aanwezig in het Devoon. De huidige paardenstaarten zijn in de loop van de aardgeschiedenis onveranderde levende fossielen.
De naaktzadigen vertonen secundaire diktegroei door tracheïden. De bladeren zijn naald- of schubvormig. De zaden zitten in kegels. Er zijn geen bloembekleedsels (perianth).
Naaktzadigen bestaan al in het Carboon.
De bedektzadigen hebben meeldraden en stampers omgeven door een perianth.
De zaden zijn omgeven door een vruchtwand. Er is altijd sprake van dubbele bevruchting.
De bedektzadigen zouden ontstaan zijn in de Jura of /en Krijt.
Als we gaan letten op de voortplanting en daarbij ook fossiele vertegenwoordigers van de afdelingen betrekken, ontstaat het volgende nogal ingewikkelde evolutionaire verhaal:
Bij de Phycofyta (de groene algen) komen heel diverse vormen van voortplanting voor.
Er is sprake van isogamie (er is geen onderscheid tussen zaadcellen en eicellen) of anisogamie (zaadcellen en eicellen zijn verschillend).
Na de bevruchting vindt er in de zygote meteen reductiedeling (meiose) plaats. Er ontstaan dus haploïde algendraden of doosjes (bij Diatomeeën).
Bij de Xantofyten ontstaan er zgn. oogonia en antheridia. Deze produceren respectievelijk eikernen en zaadcellen. De zygote gaat ook hier meteen over tot meiose.
Bij de meercellige, bolvormige Volvox worden vaak bewegelijke sporen gevormd.
Algendraden kunnen ook sporangiën vormen of gametangiën. De sporen zijn altijd haploïd. De gameten zijn meestal isogaam; aangegeven door + en -.
Bij de Cladophorales zijn de draden veelkernig en de plant is struikvormig. Er is sprake van generatiewisseling. De sporofyt wordt afgewisseld met de gametofyt. Vergelijkbaar met de situatie bij mossen en varens. Ook bij het overbekende zeesla (Ulva) komt generatiewisseling voor. Maar dan wel isomorfe generatiewisseling.
Bij Spirogyra (het spiraalwier) gaan twee draden tegen elkaar liggen, waarna de inhoud van de ene cel opgenomen wordt door de tegenoverliggende cel. De nu ontstane zygote deelt zich meteen door meiose.
De Charales zijn de meest ingewikkeld gebouwde wieren. Ze hebben een vertakte thallus met knopen en leden. Ze maken een antheridium en een oogonium. Vooral de laatste is groot met 1 eikern en is omgeven door een spiraal van draden. Soms is er sprake van enige "vruchtvorming" doordat naastgelegen cellen meedoen met het ontstaan van de bevruchte eikern. (de naam 'vrucht' heeft hier alles te maken met wensdenken).
Bij de Phaecofyceae (bruinwieren) zien we heteroplastische generatiewisseling en meiose bij de sporenvorming. Er is een haploïde gametofyt en een diploïde sporofyt. De sporofyt maakt bewegelijke zoösporen in de welbekende blazen (sporangiën) van bruinwieren die je overal op het strand tegenkomt. De kiemende spore vormt door reductiedeling meteen de gametofyt binnenin die blazen.
Er komen vele variaties op dit thema voor. Soms zijn er zelfs mannelijke en vrouwelijke planten.
Ook zijn er soorten zonder generatiewisseling.
Bruinwieren zijn reeds aanwezig in Siluur en Devoon.
Er zijn geen tussenstadia tussen bruinwieren en groene algen gevonden. Bruinwieren zijn van meetafaan hoogontwikkelde vormen met vertakkingen en sporangiën en gametangiën.
Bij de Rhodofyceae komt fycoerythrine voor als chromatofoor. In plaats van fucoxanthine zoals bij de bruinwieren. De generatiewisseling verloopt ook geheel anders dan bij de bruinwieren. Sporen en gameten zijn onbeweeglijk. Uit de zygote ontstaan sporogene draden die haploïde sporen vormen. Daaruit ontstaan meteen mannelijke en vrouwelijke gametofyten. Na de bevruchting ontstaan meteen weer sporen (door meiose).
Roodwieren kunnen evenmin zijn ontstaan uit groene algen. Vanwege bezit van andere kleurstoffen en andere vormen van generatiewisseling.
Dan komt weer de oude suggestie: wellicht ontstaan uit Cyanofyten?
Onmogelijk! Cyanofyten hebben geen kern en niet dezelfde kleurstoffen.
De voortplanting (met generatiewisseling) bij mossen en varens en paardenstaarten is hierboven al behandeld.
Als startpunt voor het ontstaan van mossen worden vaak de levermossen (Marchantiales).
Met 8500 soorten, onder vaak extreme omstandigheden groeiend en bloeiend met vlezig thalli vlak tegen de grond, zouden toch vele fossielen gevonden moeten zijn als v.b. van eerste landplanten.
Echter: de bouw van het plantje is vrij ingewikkeld en de voortplantingsorganen nog veel ingewikkelder.
https://web.archive.org/web/20050323230406im_/http://home.manhattan.edu/~frances.cardillo/plants/nonvas/marchan1.gif
https://web.archive.org/web/20050312083102im_/http://home.manhattan.edu/~frances.cardillo/plants/nonvas/marchan6.gif
Het zal duidelijk zijn dat uit levermossen niet zo gemakkelijk gewone mossen ontstaan. Er is bovendien geen enkel fossiel bewijs!
Bij mossen en varens zijn de gametofyten heel verschillend gebouwd. Die groepen kunnen dus niet uit elkaar zijn ontstaan. De sporofyten komen ook in niets met elkaar overeen.
Gametofyten bij varens en paardenstaarten vertonen wel overeenkomsten maar de sporofyten lijken in niets op elkaar. Er is daarom ijverig gezocht naar fossielen die als overbrugging tussen de grote
afdelingen varens, paardenstaarten, naaktzadigen en bedektzadigen zouden kunnen optreden.
Maar helaas!!
Veel aannemelijker is dat alle bovengenoemde 1-celligenj, schimmels, sporenplanten en zaadplanten de (ondiepe) zeeën, meren en kust- en binnenlandgebieden van Pangea bewoonden. Samen met alle hoofdafdelingen van het dierenrijk en mensen.
2.8 Fossielen van de eerdergenoemde groepen.
2.8.1 Op grond van fossiele vondsten wordt de afdeling varens teruggekoppeld aan de:
1e klasse: Psilofyten = Psilopsita.
Met als beroemd v.b. Rhynia. Rhynia heeft geen bladeren, geen wortels en de stengels zijn dichotoom vertakt. Een andere soort heeft wel zijtakjes aan een doorlopende stengel.
Rhynia heeft wel vaatbundeltjes en eindstandige sporangiën. Er zijn echter nog geen bastvaten. De houtvaten bestaan enkel uit tracheïden.
Ook komen er huidmondjes (stomata)voor op de stengeltjes.
De uiteinden van de stengels zijn soms verbreed. Zou dit een eerste vorm van bladvorming kunnen zijn?
Asteroxylon heeft wel bladschubben (microfyllen) en ook bastvaten. De vaatbundeltjes vertakken zich richting de bladschubben.
Suggestie: Rhynia zou ontstaan kunnen zijn door reductie uit Asteroxylon!
Nu nog voorkomend: de orde Psilotales. Met schubvormige blaadjes, geen wortels, generatiewisseling.
Ze groeien nu nog in tropische omgeving, dicht bij water.
https://upload.wikimedia.org/wikipedia/commons/thumb/b/bb/Starr_030222-0009_Psilotum_nudum.jpg/260px-Starr_030222-0009_Psilotum_nudum.jpg
Er zijn geen fossielen van bekend.
Conclusie: de oorsprong van Psilofyten is nog onbekend.
2.8.2 Lycopodiinae (wolfsklauwen).
Er is nog geen secundaire diktegroei. De planten hebben wel worteltjes en bastvaten (floëem).
Sporangiën staan in de bladoksels.
De prothallia leven als knolletjes ondergronds. Voor de voeding zijn ze afhankelijk van mycorrhiza (schimmeldraden). Ze zijn pas geslachtsrijp na 12-15 jr. De levensduur van de prothallia kan 20 jr. bedragen.
Fossiele Lycopodiinae komen voor in onder-Devoon. Ze zijn 300 miljoen jaar lang onveranderd gebleven. Het waren de voorlopers van Sigillaria en Lepidodendron (?)
Sigillaria.
https://upload.wikimedia.org/wikipedia/commons/thumb/e/e0/Sigillaria.png/150px-Sigillaria.png
De orde Selaginellares zijn de mosvarens. Ze leven nu met wel 700 soorten vaak als klimplanten in de tropen. Ze komen verder bijna overal op aarde voor.
De blaadjes hebben zowel spons- als pallissadenparenchym. Ze vormen microsporangiën en
megasporangiën. De antheridiën die zich ontwikkelen in de microsporangiën blijven op hun plaats zitten en produceren de spermatozoïden. Binnen een megasporangium ontstaat het meercellig prothallium met archegonium.
Selaginella spec. Er zijn wel 700 soorten.
https://upload.wikimedia.org/wikipedia/commons/thumb/0/05/Selaginella_lepidophylla_gruen.jpeg/266px-Selaginella_lepidophylla_gruen.jpeg
De orde Lepidodendrales kwamen voor in het Carboon en groeiden uit tot 40 m. hoge bomen met stammen van 5 m. dik. De bladeren waren microfyllen en het cambium vormde weinig hout maar wel een dikke kurkschors.
Lepidodendron.
https://upload.wikimedia.org/wikipedia/commons/thumb/b/b8/Stigmaria_Heimans.jpg/266px-Stigmaria_Heimans.jpg
De familie Sigillariaceae had 1 m. lange bladeren die 10 cm. breed waren. Aan de toppen van de bomen stonden grote sporofylkegels. Ze waren net als Selaginella heterospoor.
In het Carboon kwamen Lepidosperma voor. Ook wel genoemd zaadvarens omdat ze "zaden" vormden.
De megasporenwand vormde samen met de integument een 'zaadhuid".
Zo ontstonden 'zaadkegels', vergelijkbaar met de Naaktzadigen.
Bij de orde Isoetales was de ontwikkeling vergelijkbaar met die bij Selaginella.
Twee soorten komen nu nog voor in ons land: de grote en de kleine biesvaren.
https://upload.wikimedia.org/wikipedia/commons/thumb/0/02/Isoetes_tegetiformans.jpg/450px-Isoetes_tegetiformans.jpg
De prothallia zijn uiterst klein en de microprothallia ontwikkelen zich binnen de sporenwand.
Hoewel ze uiterst primitief lijken stammen de eerste isoetessoorten uit het Carboon.
Ze komen nu verspreid over de hele aarde voor en groeien in moerassige gebieden en mangrovebossen.
Zijn het sterk gereduceerde planten vanwege terugkeer naar water??
2.8.3 Equisetinae (paardenstaarten).
Ze komen al voor in het onder-Devoon.
De Hyenales hebben nog wel eindstandige sporangiën.
Calamites waren o.a. bomen van 30 m. hoog en 1 m. dik die leefden in het Carboon.
https://upload.wikimedia.org/wikipedia/commons/thumb/6/62/Calamites_reconstrucci%C3%B3n.png/185px-Calamites_reconstrucci%C3%B3n.png
De huidige kruidachtige paardenstaarten worden beschouwd als relicten van de bomen in het Carboon.
Het zijn gedurende 400 miljoen jaren onveranderde levende fossielen. Vanaf het Devoon!
2.8.4 Filicinae (varens).
Bij de orde Primofilices komen nog eindstandige sporangiën voor. Ze worden beschouwd als tussenvorm van varens en paardenstaarten.
Bij Cladoxylon zijn de blaadjes nog erg klein en de sporangiën zitten aan de bladranden.
Bij de Lepidospermae en de Pteridospermae blijft de megaspore binnen het sporangium en het integument verbonden met de sporofyt. Dit lijkt op begin van zaadvorming. Ze worden daarom ook wel gymnospermopsida genoemd. Maar die vorm van zaadvorming kwam al voor in het Carboon, voordat de meeste varens verschijnen. Dan kan het geen eindpunt van een ontwikkelingsreeks zijn.
Misschien: varens ontstonden parallel met mossen uit de groene algen. Niemand weet hoe!!
2.8.5 Onderafdeling Gymnospermen (naaktzadigen).
1e klasse: Pteridospermae; zie boven.
2e klasse: Cycadinae. Ze leven nu vooral in de tropen. Het vruchtblad toont een steeds verdere reductie. Tenslotte zijn er nog 2 zaden per vruchtblad. Ze kwamen al voor in het Perm. Vele soorten nu als kamerplant gekocht.
https://upload.wikimedia.org/wikipedia/commons/thumb/2/2a/Sorakuen03s3200.jpg/266px-Sorakuen03s3200.jpg
3e klasse: Benettitinae.
Ze zouden stammen uit het jonge Paleozoïcum (Devoon). Ze hebben een aanleg van een perianth (bloembekleedsel). Daarmee zouden ze tot de vroege angiospermen gerekend kunnen worden.
Er zou al insectenbestuiving hebben plaats gevonden.
https://upload.wikimedia.org/wikipedia/commons/thumb/4/46/Bennettitales-cycadeoidaceae.jpg/220px-Bennettitales-cycadeoidaceae.jpg
4e klasse: Cordaïtinae. Fossielen vanuit Carboon/Perm.
Er is zowel sponsparenchym als pallissadenparenchym aanwezig. Eveneens secundaire diktegroei (vanuit cambium). De orde Cordaïtales zouden de voorvaders zijn van de Coniferen en Ginkgo's.
Ook bij de Cordaïtales ontstaan de sporen in aparte mannelijke en vrouwelijke kegels. Zaden en kegels zijn echter heel anders gebouwd dan bij de coniferen. Zijn na het Perm de Coniferen en Ginkgo's ontstaan via de Cordaïtales? Coniferen en Ginkgo's zijn de dominerende plantengroepen in het Trias, Jura en Krijt. Daarom ook op Pangea? Zie de pg. 'Een Bijbels georiënteerde geologie'.
5e klasse: Ginkgoinae. Ze zijn afkomstig uit Carboon/Perm en komen nu nog alom op aarde voor. Ginkgo (Japanse notenboom) veel als sierbomen aangeplant, ook in ons land.
https://upload.wikimedia.org/wikipedia/commons/thumb/4/45/Ginkgo_biloba_JPG1a.jpg/100px-Ginkgo_biloba_JPG1a.jpg
6e klasse: Coniferae.
De fossiele soorten Ephedra, Welwitschia en Gnetum groeiden in drogere gebieden in b.v. Australië in de tijd van het Tertiair. Vond er ontwikkeling plaats in de richting van de Angiospermen ?
Dit zou blijken uit het bezit van tracheeën, netnervatuur, perianth, insectenbestuiving en geringere ontwikkeling van antheridium en archegonium.
Of groeiden ze al op drogere gebieden in de binnenlanden van Pangea, samen met angiospermen?
De klassen van de Gymnospermen vertonen zeer lange gescheiden "ontwikkelingslijnen". In het Devoon kwamen ze gelijktijdig voor met Pteridospermen (zaadvarens) en Eusporangiate varens.
Misschien toch ontstaan uit Psilotumachtige planten??
Of ook mogelijk: Coniferen gaan terug op Lycopodiumsoorten (wolfsklauwen)??
2.8.6 Onderafdeling: Angiospermae (bedektzadigen).
Bestuiving door wind, insecten, vogels, zoogdieren of water.
De stuifmeelkorrels ontkiemen op de stempel van dezelfde soort plant. Er groeit een stuifmeelbuis door de soms heel lange stijl naar de zaadknop. De zaadknop met de embryozak is te vergelijken met het archegonium van de sporenplanten. Zo is ook de stuifmeelbuis te vergelijken met het antheridium van de sporenplanten.
De beide gametofyten zijn dus steeds verder gereduceerd (?)
De sporofyten zijn op vele manieren totaal verschillend gebouwd.
Ontwikkelingslijnen binnen de vele orden en families van de angiospermen zijn nauwelijks te trekken.
Wel wordt de schroefvormige stand van de bloemdelen als oorspronkelijk beschouwd. Dit komt voor bij b.v. Ranunculaceae en Magnoliaceae. Vaak is er wel uitval van een 'ring'.
Vruchtvorming vindt op totaal verschillende manieren plaats. Vanuit vruchtbeginsel, bloembodem en/of andere bloembekleedsels.
Bij insectenbloemen zien we vaak een dorsiventrale bouw van de bloem. B.v. lipbloemigen, vlinderbloemigen etc. Maar die komt ook bij grassen voor.
Bij Nymphaea is een opvallende overgang tussen kroonbladen en meeldraden te zien.
Op grond hiervan worden verregaande conclusies getrokken over het ontstaan van meeldraden uit bladeren.
Er is soms sprake van het voorkomen van pseudanthiën. Het zijn oneigenlijke (valse), op bloemkroon- bladeren gelijkende, schutbladeren zoals bij de Araceae.
Soms zijn meeldraden gereduceerd (?) tot nectariën (honingklieren).
Er komen vele varianten van bovenstandig, onderstandig en halfonderstandige vruchtbeginsels voor.
Als oorspronkelijk (dus als eerder voorkomend in een evolutielijn) gelden:
1. een onbestemd en vaak groot aantal bloemdelen
2. schroefvormige of radiale plaatsing van bloemdelen tgo. verspreide plaatsing
3. een 2- of 3-tallig bloemdek
4. vrijstaande bloembladeren tgo. vergroeide
5. apokarpe gynaecium tgo. syncarpe (vergroeide vruchtbeginsels)
6. radiaire bloemen tgo. symmetrische
7. gelijkvormige tgo. ongelijkvormige bloemen
8. bovenstandig tgo. onderstandig en halfonderstandig vruchtbeginsel
9. zaadknoppen met 2 integumenten
10. zaad met goed ontwikkeld endosperm (reservevoedsel)
11. atrope (rechtopstaande) zaadknop tgo. anatrope en campylotrope zaadknoppen
12. 2-kernige tgo. 3-kernige stuifmeelbuis
13. ontbreken van tracheeën in het secundaire hout.
Als duidelijk afgeleid gelden: het voorkomen van functieloze organen, extreme aanpassingen, parasitaire planten, carnivore planten (zoals zonnedauw en bekerplanten) en 1-jarige planten.
Vaak leveren bovenstaande kenmerken tegenstrijdige conclusies op. Sommige onderdelen van een plant kunnen hoogontwikkeld zijn terwijl andere weinig ontwikkeld lijken. Het onderscheid tussen 1- en 2-slachtige bloemen, 1-huizige of 2-huizige planten, windbloemen en insectenbloemen is in dit opzicht niet eenduidig.
Het blijft moeilijk om tot een natuurlijk systeem van opeenvolgende plantengroepen te komen.
De oudste fossielen van Angiospermen stammen vooral uit het onder- Krijt, gevonden in Noord-Amerika en Groenland. Het zijn merkwaardigerwijs vertegenwoordigers van Policarpicae (zoals Magnoliaceae, Monochlamideae (b.v. Fagaceae (beuken) en Moraceae (vijgen en moerbeien) en Monocotylae (éénzaadlobbigen zoals grassen).
De Angiospermen stammen dus al uit de Jura. Ze zouden daarna de eerder genoemde Benettitinae verdrongen hebben.
De welbekende Kardinaalsmuts (Euonymus europeus) kwam al voor in het vroeg-Krijt. De kardinaalsmuts behoort tot de familie Celastraceae.
De website 'Encyclopedia.com' vermeldt:
Celastrophyllum circinerve One of the earliest flowering plants known to palaeobotanists, belonging to the family Celastraceae which pre-dates most others. The family began with C. circinerve in the Early Cretaceous and survives today in the form of Euonymus europaeus, the spindle tree (kardinaalsmuts).
Opm.: Volgens het nieuwe Cladistische systeem zijn de 1-celligen verdeeld over alle domeinen van de organismen:
Unikonta
Excavata
Archaeplastida
Rhizaria
Chromalveolata
Hacrobia.
Basidiomyceten en Ascomyceten, Amoebozoa, insecten, zoogdieren en mensen zijn samen onderdeel van de Unikonta.
Euglena en Trypanosoma behoren tot de Excavata.
Roodwieren, groenwieren, mossen, varens en bloemplanten zijn onderdeel van de Archaeplastida.
Foraminiferen en Radiolariën behoren tot de Rhizaria.
Pantoffeldiertjes, Dinophyta, Oomycetes (o.a. Phytophtora), Bruinwieren, Diatomeeën etc. vormen samen de Chromalveolata.
De Hacrobia zijn de Haptophyta, Cryptophyta en de piepkleine Picozoa.
2.9 Voorlopige beoordeling van evolutionaire ontwikkelingen.
Voor de beoordeling van de veronderstelde evolutionaire ontwikkelingen van bacteriën, schimmels algen en sporenplanten en zaadplanten zou ik willen uitgaan van de volgende uitgangspunten:
2.9.1. Verreweg de meeste fossielen, zowel van planten als dieren, worden gevonden in ondiepe zeeën, in riviermondingen, in estuaria of lagunes, rond meren en moerasgebieden. Ook levend op koraalriffen die zich in niet al te diepe kustwateren konden ontwikkelen. Daarom is het vrij logisch om de aardperioden Precambrium, Cambrium, Ordovicium, Siluur, Devoon, Carboon en Perm samen te voegen tot één aardperiode.
2.9.2. Geologen onderscheiden 5 extinctiperioden in de aardgeschiedenis. Het betekende het uitsterven van 80 à 95 % van alle soorten organismen. Het wil ook zeggen dat er in de loop van de aardgeschiedenis wel 10, 100 of zelfs 1000 keer zoveel soorten organismen hebben geleefd dan er nu op aarde zijn. Het is daarom niet vreemd dat vele fossiele plant- en diersoorten een tussenpositie innemen binnen de huidige soorten. Deze zgn. "tussenvormen" of "overgangsvormen" zijn op zich genomen daarom geen enkel bewijs voor evolutie van nieuwe soorten of geslachten.
2.9.3. Het lijkt me logisch om bij elke periode in de aardgeschiedenis te beseffen dat er tegelijkertijd heel verschillende leefgebieden voorkwamen: ondiepe zeeën, rivieren, waddengebieden, stranden en duingebieden, moerassen, bossen, regenwouden, savannen (steppen), graslanden, heuvels en
bergen .............. Wellicht zelfs ijzige poolgebieden en kleine woestijnen.
Bezet met talloze, totaal verschillende dier- en plantengroepen. Waarbij zgn. hoogontwikkelde organismen de eenvoudiger gebouwde organismen niet verdringen of zelfs tot uitroeiing dwingen. Dat gebeurt in de huidige wereld evenmin! Bij een groot aantal niches kunnen (bijna) alle vroegere en nu levende soorten naast elkaar bestaan.
Als we alle gevonden fossielen zouden groeperen naar hun diverse leefgebieden en daarbij niet bij voorbaat koppelen aan miljarden of miljoenen jaren geleden zou er een heel ander "plaatje" tevoorschijn kunnen komen.
2.9.4. Het ontstaan van wel tot 100 m. dikke steenkoollagen, biljoenen liters aardolie en aardgas, enorme bruinkoolvelden en vooral de enorme zoutvelden onder West-Europa en vele andere werelddelen is nog steeds volledig onverklaarbaar. Bedenk dat er b.v. 10 m. plantenmateriaal noodzakelijk is om 1 m. steenkool te vormen. Hoeveel er nodig is voor alle aardolie en aardgas? Geen idee!
Plant- en dierlijk materiaal vrij langzaam overdekken met sediment biedt geen enkele verklaring.
Dat geldt ook voor de meters dikke zoutlagen met hoge pieken, tot duizenden meters; zowel onder ons land en op vele andere plaatsen in de wereld. Een zeebekken waarin vrij langzaam water verdampt kan nooit de oorzaak zijn.
2.9.5. Gebergtevorming tijdens de continentverschuiving waarbij de huidige ligging van de werelddelen werd bepaald kan oorzaak geweest zijn van ongelofelijke opeenhopingen van fossiele dieren en planten.
Gedeeltelijk kan ze ook de "opeenvolging" van dierlijk en plantaardig leven wellicht verklaren.
Er worden eveneens 5 perioden van orogenese onderscheiden. Hooggebergten die tijdens de laatste orogenese ontstonden waren de Alpen, de Andes, de Rocky Mountains, de Himalaya en misschien ook de Kilimanjaro en de Ararat.
Opm. In het boek Earth History and Palaeogeography worden zelfs 48 perioden van orogenese benoemd.
Deze periodes kunnen van vrij korte duur geweest zijn (enkele honderden of duizenden jaren).
Tijdens een eerdere periode van gebergtevorming als gevolg van continentverplaatsing kunnen de gebergtegebieden zoals de Ardennen, de Schotse Hooglanden, de Pyreneeën, de Appalachen, de Noorse berggebieden enz. zijn ontstaan. Ook met grote gevolgen voor de temperatuurverdeling over de landstreken, het ontstaan van specifieke groepen fossielen etc.
Hoofdstuk 3 De opeenvolgende geologische perioden.
Het volgende is een poging om een deel van de gegevens die bekend zijn van omstandigheden (o.a. milieu) en fossielen van de aardlagen in kaart te brengen.
De boeken die ik hiervoor tot mijn beschikking heb zijn de volgende:
1. Het al eerder genoemde Lehrbuch der Botanik, Fisher Verlag, Stuttgart. 1967
2. Mossen en Vaatplanten, Oosthoek's uitg., door Dr. C. Kalkman 1972
3. The Genesis Flood, uitg. Baker Bookhouse, Grand Rapids 1965
4. The Biblical Flood and the Ice Epoch, uitg. Pac. Meridian Publ.l Co., Seatle 1966
5. Ook eerder genoemd: Les Iguanodons de Bernissart, door Edgard Casier. 1960
6. Opkomst en ondergang van de Dinosaurus, uitg. Ambo, door Steve Brusatte 2018
7. Encyclopedia of Dinosaurs, uitg. Beekman House, New York, door vele geleerden, kunstenaars en
fotografen. 1990
8. De prehistorische wereld, uitg. Hamlyn Publ. group te Londen Ned. vert. door uitg. icob
door Richard Moody 1980
9. Evolutie, een historisch panorama, uitgeg. door Tirion natuur, door Douglas Palmer 2009
10. Oorsprong, wetenschap[ en bijbel verenigd, uitg. Groen, door Ariel Roth 2005
11. Terug naar de oorsprong, uitg. De Oude Wereld, door Peter Borger 2009
12. Evolutie, het nieuwe studieboek, uitg. De Oude Wereld, door Reinhard Junker en Siegfried
Scherer. 2010.
13. Contested Bones, uitg. FMS Publications, door Christopher Rupe en John Sanford, 2019
14. Het ontstaan van soorten, uitg. Olympus (Atlas Contact), door Charles Darwin 1859/2010
15. Het verhaal van onze voorouder, een pelgrimstocht, uitg. Nieuw Amsterdam, door Richard
Dawkins en Yan Wong, 2017.
16. Earth in upheaval, uitg. Victor Gollansz en Sidgwick and Jackson, London. 1956
17. Biodiversity and Earth History, uitg. Springer- Verlag Berlin Heidelberg, door Jens Boenigk,
Sabina Wodniok en Edvart Glücksman. 2015.
18. Earth History and Palaeogeography, uitg. Cambridge University Press, door Trond H. Torsvik en
L. Robin M. Cocks. 2017 derde druk 2019.
3.1 De eerste periode (precambrium en cambrium t/m Perm).
Aan de rotsige kusten van West- Australië worden de Stromatolieten gevonden. Ze kwamen over de hele wereld voor. Het zijn samenballingen van algen en cyanobacteriën. Ze vormen bolvormige oppervlakteklitten in sediment van kiezel, kalksteen en hoornsteen. Het eerste leven, 3.5 miljard jr. geleden? Het tijdperk wordt Archaea genoemd.
Daarnaast worden er vondsten gemeld van 2 miljard jr. geleden. Gevonden in hetzelfde gebied en in Rusland en Canada. Dan gaat het over 'bacterieachtige filamenten' van cyanobacteriën gevonden in kiezelhoudend hoornsteen. In de omgeving van vulkanische bronnen. Critici beweren dat het slechts stapels minerale korrels zijn.
Er wordt verondersteld dat er vanaf 640 miljoen jaar geleden enorme ijstijden zijn geweest. Met gemiddelde jaartemperaturen van -500 C. De oceanen waren toen zelfs bevroren. Als bewijs daarvoor worden het voorkomen van zgn. valstenen genoemde, samen met krassen in de rotsen. Of dat voldoende bewijs is?!
De eerste sponzen kwamen, denkt men, al 700 milj. jr. geleden voor.
Sponzen zouden de enige Metazoa (meercellige dieren) zijn die rechtstreeks van 1-celligen zouden af stammen. Terwijl dit toch minder simpel is dan het lijkt want binnen een spons is er al een grote taakverdeling tussen 'skeletcellen' met zgn. spiculae, zenuwcellen en kraagcellen met zweephaar die zorgen voor een waterstroom om voedingsstoffen naar de cel brengen. Daarnaast zijn er nog
- sluitcellen die de waterstroom mogelijk maken
- contractiele cellen die de waterstroom kunnen regelen
- dekcellen die de huid vormen
- amoebocyten zorgen voor transport van voedingsstoffen,
vorming van skeletnaalden en van voortplantingscellen
- archeocyten die kunnen uitgroeien tot eicellen.
Over de evolutie van sponzen is niets met enige zekerheid bekend.
3.1.1 Cambrium.
Na een aantal veronderstelde wereldomvattende ijstijden volgt de Cambrische explosie van leven.
Leven vanaf 570-540 miljoen jr. geleden? Voorafgegaan door de Ediacara-fauna.
Deze fauna is bedekt door fijnkorrelig zand en modder. In lagen van 3 km. dikke vulkanische as vond men 30 verschillende soorten dieren met enkel zachte weefsels; lijkend op kwallen, ringwormen en weekdieren. Allemaal bodemdieren?
Het zijn hoogontwikkelde organismen die kunnen waarnemen, voortbewegen, zich voeden en voorplanten.
In het zgn. Cambrium worden dan langs dezelfde kusten in ondiepe zeeën schelpdieren, weekdieren, sponzen, armpotigen (b.v Longtancunella, een brachiopode, zoals de nu levende Nautilus) en arthropoden (geleedpotigen) gevonden. Ze leven op riffen.
Darwin schreef al: "de toenmalige wereld wemelde van de levende wezens". Het was een groot probleem voor zijn theorie!
Vooral de
Anomalocaris is een vrij grote, snel zwemmende rover met grote ogen en vele
 Anomalocaris. bron Wikipedia.
Anomalocaris. bron Wikipedia.
"zwemvliezen". Myllenkunmingia is een chordaat, dus een voorloper van alle gewervelden.
Het snel verschijnen van talloze groepen die vaak ver ontwikkeld zijn met skeletten verbijsterden de paleontologen. Ze wordt nog steeds niet begrepen. Ze worden al gevonden in het onder-Cambrium. En dan moet er snelle afdekking hebben plaats gevonden in een milieu zonder aaseters en bacteriën zegt Richard Moody in zijn boek De prehistorische wereld. Wanneer zou zo'n wereld ooit hebben bestaan?
In zijn boek geeft Moody een eenvoudig schema van ontstaan van het leven der dieren.
Vanuit het precambrium ontstaan:
- blauwgroene algen
- eencellige dieren
- sponzen
- holtedieren
- wormen
Vanuit het cambrium ontstaan:
- brachiopoda (armpotigen)
- mosdiertjes
- amphineura (chitons)
- gastropoda (slakken)
- koppotigen (inktvissen)
- stekelhuidigen (zeesterren etc.)
- trilobieten (geleedpotigen)
- graptolieten
- nautiloïden (vgl. inktvissen)
- peripatus (fluweelworm)
- gewervelde dieren.
aar kwamen al die complex gebouwde dieren vandaan? Ze kwamen voor over de hele wereld!
M.b.v. de zgn. moleculaire klok (die uitgaat van veranderingen in macromoleculen door willekeurige mutaties) zouden deze Ediacara-wezens al 400 miljoen jr. eerder ontstaan moeten zijn.
De Burgess Shale in Canada behoort tot één van de rijkste fossiele vindplaatsen van de "eerste wereld".
Sinds de laatste onderzoekingen worden de daar gevonden fossielen beschouwd als de eerste vertegenwoordigers van alle nu nog levende groepen. Vooral beroemd is Pikaia, misschien een voorloper van de chordata.
Tijdens de Cambrische explosie zijn de meeste grote groepen dieren (de stammen of fyla) ontstaan: sponsen, zeeanemonen, trilobieten en wormen tot en met de vertebraten (gewervelde dieren, met chorda en inwendig (kraakbeen)skelet).
Een volgende bewering komen we bij evolutionisten vaak tegen: "door een continu evolutionair verloop van soorten en families zijn de cambrische soorten uitgestorven". Dit is een flinterdunne, vrijwel nietszeggende redenering. Er moeten concrete oorzaken zijn voor het uitsterven van organismen. Als je die niet weet moet je ernaar gaan zoeken. Er leven nu op aarde miljoenen soorten naast elkaar die elkaar bijna nooit verdringen of uitroeien! Wel bezetten ze verschillende 'niches'.
3.1.2 Ordovicium.
In het zgn. Ordovicium leven dieren van dezelfde diergroepen op dezelfde plaatsen als tijdens het cambrium.. Alleen de "continenten" zijn verder aan de wandel gegaan. Prachtig gebouwde kreeften, trilobieten, brachiopoden, arthropoden en zeelelies (Crinoidea) komen voor. En vooral: pijlinktvissen. Wel zo ongeveer de meest prachtig gebouwde "roofvissen" op aarde. Met ogen die vergelijkbaar zijn qua bouw met onze ogen.
Graptolieten (al aanwezig vanaf Ediacara) en Ostracoden (mosselkreeftjes, wel 65.000 soorten fossiel bewaard) zijn uitbundig voorkomende organismen in deze tijden.
Samen met de Agnatha (tandeloze vissen) en Linguliden; levende fossielen.
https://upload.wikimedia.org/wikipedia/commons/thumb/3/3b/Lingula-Ozamis-2.JPG/1280px-
Lingula-Ozamis-2.JPG
Vindplaats van deze fossielen: een dorpje hoog in het Andesgebergte. Maar dat was oorspronkelijk een ondiepe zee!
Ook
Conodonten (een palingachtige vis met echte vinnen) komen voor.
 Een Conodont. bron Wikipedia.
Een Conodont. bron Wikipedia.
Daarnaast verschijnen vissen met tanden en grote roofzuchtige kreeftachtige arthropoden. De Eurypterida met grote ogen en grote scharen worden wel 2 m. lang.
 Pterygotus, een Eurypteride. Jagend op vissen. bron Wikipedia.
Pterygotus, een Eurypteride. Jagend op vissen. bron Wikipedia.
3.1.3. Siluur.
Tijdens het Siluur blijft het verhaal ongeveer hetzelfde: veel riffen in ondiepe zeeën met prachtige trilobieten, zeelelies, koralen en schelpdieren. Zwaar gepantserde Agnatha komen samen voor met niet gepantserde snel zwemmende vissen. En krabben, kreeften, degenkrabben (Limulidae), schorpioenen en Eurypterida (grote waterschorpioenen).
Aan het eind van die periode wordt het interessanter omdat de eerste landplanten in die afzettingen worden gevonden in de moddervlakten langs de kust: de bladloze sporenplanten Cooksonia (dichotoom vertakte stengel) en Steganotheca (met vertakkingen van de 1e en 2e orde). Samen met mossen en korstmossen.
Dit zegt weinig tot niets over plantenleven in overige landstreken van het enorm grote Pangea.
Daarbij een spinachtige land-arthropode, de Degenkrab, ook een levend fossiel.
https://commons.wikimedia.org/wiki/File:Mangrove_Horseshoe_Crab_(Carcinoscorpius_rotundicauda)_(6707310189).jpg
3.1.4 Devoon.
In het Devoon (400 miljoen jr. geleden?) zijn er afzettingen in kiezel en organisch afval. In de buurt van hete bronnen op het land. Er komen enkele nieuwe plantensoorten, o.a. de bladerloze Rhynia die ook nu nog alom op aarde voorkomt. Maar ook Asteroxylon met schubvormige bladeren. Varens, paardenstaarten en wolfsklauwen beginnen aan hun opmars. Dus alleen sporenplanten.
Het zeeleven (nu tot wel 100 m. diep) wordt gevarieerder: haaien die al in het Siluur waren ontstaan floreren nu, samen met echte beenvissen en rogachtige vissen. En bovenal een Sarcopterygische (behorend tot de kwastvinnigen) longvis.
Zowel inktvissen, haaien en kwastvinnigen doen qua lichaamsstructuren in niets onder voor die van zg. "hogere" waterdieren.
De dieren van het zeeleven zijn nu:
sponzen, koralen, brachiopoden, crustaceeën, echinodermen, arthropoden, inktvissen en zeespinnen.
De leiplaten in Duitsland bevatten vele schitterende fossielen; dus ontstaan door bedekking met modder. In de leisteen van Australië vindt men Sarcopterygii (kwastvinnigen) en actinopterygii (echte beenvissen, de straalvinnigen). Ook vele Placodermi (zwaar bepantserde bodemvissen).
Er is daarom geen zwaarwegend argument om de moderne vissen te laten ontstaan uit die laatstgenoemde bodemvissen.
Uit de kwastvinnigen zouden de landdieren moeten zijn ontstaan omdat de borst- en buikvinnen beenachtige structuren bevatten die lijken op de botten in de ledematen van de amfibieën enz.
https://upload.wikimedia.org/wikipedia/commons/thumb/4/4b/Coelacanth.png/266px-Coelacanth.png
Maar de moderne coelacanth (Latimeria) leeft nog steeds in de diepzee, voor de kust van de Comoren, in de Indische Oceaan. Wel heel vreemd dat ook de rugvin de bouw van een toekomstige looppoot vertoont!
In Canada werden in hetzelfde type kustafzettingen duizenden fossielen verzameld. Vooral
Eustenopteron werd wereldberoemd omdat ook wij daaruit ouden zijn ontstaan.
 Eustenopteron. bron Wikipedia.
Eustenopteron. bron Wikipedia.
De modder in de riviermonding was vermengd met planten. O.a. Archaeopteris, met varenachtige bladeren en een houten stam tot 18 m. hoog. Van het gymnosperme type. Maar zonder zaden.
Deze boom leverde in ieder geval een overvloed van voedsel voor de waterdieren.
Fossielen van eenzelfde soort gebied, nu onder het ijs gevangen in het noorden van Canada, bewees het bestaan van Tiktaalik. Een wonder van een overgangstype van kwastvinnige naar een tetrapode (een viervoetig landdier)!? Het heeft een onderling verbonden schedel met nek, "schouders" ('borststreek'), "armen" en de "gewrichten" van 'elleboog' en 'pols'.
Tiktaalik.
https://upload.wikimedia.org/wikipedia/commons/thumb/2/2b/Tiktaalik_roseae_life_restor.jpg/266px-Tiktaalik_roseae_life_restor.jpg
Enthousiast was men eerst over de vondsten van
Ichtyostega (een salamanderachtige vis) die m.b.v. 5-tenige poten het land op kon. Echter: het dier had een grote 'visse'-staart!
 Acanthostega. bron Wikipedia.
Acanthostega. bron Wikipedia.
En Acanthostega maakte duidelijk dat de ledematen (met ongeveer 8 tenen) vooral goed waren voor het waterleven. Ze hadden meerdere tenen en ademden m.b.v. kieuwen.
3.1.5 Carboon.
Grote equatoriale zeeën tot 40 m. diep en vele meren met hete bronnen en omringende bossen werden veelal bedekt door vulkanische as.
Vanaf deze periode vindt de vorming van enorme steenkoollagen plaats.
De eerste periode, volgens de geologische tijdschaal, waarin het landleven uitbundig aan bod komt,
Honderden soorten insecten bevolken gebieden rond meren, waaronder hooiwagens. Ook schorpioenen met boeklongen (net als bij spinnen) zijn aanwezig. Belangrijke amfibie is Balanerpeton.
Balenerpeton, het eerste viervoetige landdier?
https://upload.wikimedia.org/wikipedia/commons/thumb/d/d3/Balanerpeton_BW.jpg/290px-Balanerpeton_BW.jpg
Balanerpeton werd ontdekt in Schotland. Wel 30 dieren bij elkaar. Het dier heeft gehoorbeentjes, maar legt nog eitjes in het water.
In een marine lagune in de VS werden wel 108 soorten (4800 stuks) vissen ontdekt, waaronder haaien, beenvissen en roggen die normaal niet fossiliseren. Daarbij vele soorten wormen met een week lichaam. Samen dus snel begraven.
In de moerassen rond de estuaria van de rivieren, meestal langs vaste kusten, groeiden boomhoge wolfsklauwen, zaadvarens, Cordaïtes, Calamites, Sigillaria en paardenstaarten. Glossopteriden (o.a. Austroglossa) kwamen voor over de hele wereld. Het zijn bomen met bladeren waarop mannelijke of vrouwelijke voortplantingsorganen staan. De vruchtlichamen die gevormd worden bevatten zaden. Samen met 150 soorten arthropoden zoals libellen, 1000-potigen, kakkerlakken, schorpioenen en amfibieën. Het boek 'Tirion Natuur' schrijft dan: "Bij overstromingen werden planten en dieren meegetrokken naar estuariene wateren, waar hun restanten werden afgezet naast brakwater- en zeedieren zoals kwallen, vissen, schelpdieren en het vreemde wezen Tullimonstrum (het nationaal symbool van Illinois).
Tullimonstrum bezat geen harde delen.
https://upload.wikimedia.org/wikipedia/commons/thumb/6/67/Tullimonstrum_NT_small.jpg/260px-Tullimonstrum_NT_small.jpg
Regenwouden groeiden ook in moerasgebieden rond een binnenzee in Nova Scotia. Op drogere gebieden groeiden naalden dragende struiken. Steeds wisselende zeeniveaus zouden gezorgd hebben voor meer dan 900 m. afzetting bestaande uit 45 lagen steenkool!
Opeenhopingen van veen en lagen sedimenten wisselden elkaar af. Nieuwe plantengroei kon wortelen in de sedimentlagen. Het veen werd turf, de turf geleidelijk omgezet in bruinkool en die op haar beurt eerst in zachte koolsoorten (eierkolen) en tenslotte in de harde antraciet. Zo vermoedt men.
Er werden meer dan 148 soorten landslakken gevonden.
In Tsjechië ontstonden grote steenkoollagen met daarin vele reptielen: Anthracosauriërs, Temnospondylen en Microsauriërs. Ook de eerste synapside reptielen. Daaruit ontstonden de zoogdieren, zo meent men.
En er kwamen reptielen voor. Die moeten eieren met schalen op het land gelegd hebben.
Echte varens groeiden er.
Er zijn vele grote bosbranden geweest.
In grote kolenlagen in Tsjechië (afkomstig van afzettingen in meren) worden duizenden goed geconserveerde fossielen gevonden met vele kleine sauriërs.
Tijdens het Carboon zouden de eerste amfibieën en reptielen zich hebben moeten ontwikkeld uit de kwastvinnigen.
Die geschiedenis is echter verborgen gebleven door de transgressie van de ondiepe zeeën in het Onder-Carboon schrijft Moody. Vanwege het ontbreken van terrestrische afzettingen.
In het Boven-Carboon had de zee zich terug getrokken en ontstonden er grote delta's en moerassen langs de kusten van Noord-Amerika en Europa. Hier leefden vele families en talloze soorten Amfibieën. Sommige met reusachtige afmetingen (4.5 m.). Kleine soorten moesten zich zien te redden en werden verdreven naar de randgebieden van de moerassen. Daar ontdekten ze de mogelijkheid van eieren leggen op het land. Die moesten dan wel stevige vliezen en harde schalen bezitten. Het amniote ei! Hoe krijg je het voor elkaar! Daarvoor moet een complete inwendige ontwikkeling plaats vinden om zo'n ei te fabriceren! Tegelijkertijd maakten ze een geschubde huid. En natuurlijk ander (voortplantings)gedrag met bijbehorende voortplantingsorganen.
300 miljoen jr. geleden kwam er een ijstijd over Zuid-Amerika, Afrika en Australië vanuit Antarctica. Dit schijnt te blijken uit puin en groeven in gesteenten. Is dit voldoende bewijs??
3.1.6 Perm.
Dit is te beschouwen als de laatste periode van de eerste cyclus. Aan het einde van het Perm komt nl. een geweldig extinctie. Veroorzaakt door enorme vulkaanuitbarstingen. Vooral in Siberië.
In Australische moerassen werd zoveel steenkool gevormd dat er nu nog 350 miljoen ton per jaar kan worden afgeleverd! Een 300 m. dikke laag organisch materiaal was daarvoor nodig! Dit gebeurde in
's winters erg koude moerassen en venen, vergelijkbaar met de taiga in Siberië en Canada.
In Zuid-Afrika ontstonden grote sedimentvlakten vanuit de gebergten in Antarctica. Hier leefden de Cynodonten, waaruit de zoogdieren zouden zijn ontstaan. Procynosuchus (met zoogdierkenmerken) leefde hier samen met vele "sauriërs".
In het Wolgabekken, met vele seizoensrivieren en overstromingsgebieden leefden de dinocephalische therapsiden ("zoogdieren") samen met Gorgonopsia, Therocephalia en Cynodontia.
In Siberië werd 7 miljoen km2 overdekt met basaltlava. Dit ging samen met vulkanisme op de zeebodem en dus zeer waarschijnlijk ook met orogenese-processen. Dit had een enorme uitstoot van CH4 tot gevolg en daardoor grote temperatuurstijging.
Archosaurus (een diapside) overleeft.
In de equatoriale wateren van rivierdelta's raken de tetrapoden wijd verbreid over het ontstane Pangea.
Vanaf het Carboon kreeg die landmassa steeds meer vorm en krijgen Afrika en Zuid-Amerika duidelijker contouren. Grote reptielen dreigen amfibieën te gaan overheersen.
Edafosaurus en Dimetrodon dragen grote zeilvormige structuren op hun ruggen.
https://upload.wikimedia.org/wikipedia/commons/thumb/0/0e/Dimetrodon_skeleton.JPG/250px-Dimetrodon_skeleton.JPG
Over de functie van het rugzeil zijn de geleerden het intussen niet meer eens. Temperatuurregeling leek eerst het meest logisch. Nu denkt men eerder aan het maken van indruk bij het zoeken van een voortplantingspartner. Vgl. de pauwenstaart. Fossielen van deze diergroepen werden veel gevonden in Texas en in Duitsland. In oorspronkelijk moerassige gebieden. Op een hooggelegen vlakte in Duitsland kwamen Eudibamus en Seymouria voor. Eudibamus was een snelle renner op twee lange achterpoten.
Eudibamus.

https://upload.wikimedia.org/wikipedia/commons/thumb/a/ae/Eudibamus_BW.jpg/260px-Eudibamus_BW.jpg
Seymouria.
https://upload.wikimedia.org/wikipedia/commons/thumb/6/6e/Primitive_Cotylosaur_Seymouria._Wellcome_M0006645.jpg/185px-Primitive_Cotylosaur_Seymouria._Wellcome_M0006645.jpg
Seymouria werd lang beschouwd als de overgang (missing link) tussen amfibie en reptiel.
Het skelet vertoont reptielachtige kenmerken. Echter: naderhand bleek dat de larven echte watervisjes waren. Dus was het een amfibie! Dan wel een zeer gespecialiseerde: planteneter of vleeseter.
In Australië bestond het moerassig loofbos uit boomhoge zaadvarens (Glossopteris), coniferen, wolfsklauwen en paardenstaarten.
De Glossopteriden (zaadvarens) groeiden in alle delen van het supercontinent.
In de Grote Karoo (1500 km. breed in Zuid-Afrika) leefden vele sauriërs tussen dezelfde planten (bomen) en de cynodonten. De laatste worden geacht de voorlopers van zoogdieren te zijn. En: ook wij worden geacht cynodonten te zijn.
Cynognathus.
https://upload.wikimedia.org/wikipedia/commons/thumb/f/fd/Cynognathus_BW.jpg/266px-Cynognathus_BW.jpg
De Cynodonten ("hondtandigen") hadden een verhemelte-plaat die mond- en neusholte van elkaar scheidde. Eten en ademhalen kon dus tegelijkertijd gebeuren. De onderkaak bestond nog wel uit diverse onderdelen. Die werden later opgenomen in het middenoor als gehoorbeentjes.
Hoe dat proces ooit verlopen moet zijn? Geen bewijzen vanuit fossielenreeksen!
Door vele overstromingen, dankzij de moessonregens, verdronken de dieren en fossiliseerden.
In het Wolgabekken vond men honderden skeletten van Therapsiden (dinocephalische Therapsiden), Gorgonopsiden, Therocephalia en Cynodontia.
Arctops, een Therapside.
https://upload.wikimedia.org/wikipedia/commons/thumb/f/f1/Arctops_ferox112.jpg/220px-Arctops_ferox112.jpg
Therapsiden worden tot de Mammalia gerekend. Maar: hoe is de voortplanting??
An het einde van het Perm vond men ontsluitingen van de Siberische Trappen.
https://upload.wikimedia.org/wikipedia/commons/thumb/0/00/Extent_of_Siberian_traps_german.png/300px-Extent_of_Siberian_traps_german.png
Lagen van honderden meters dik en turflagen van 20 m. dik en een opp. van 30.000 km2. En basaltlava van 7 000 000 km2 Samen met de ontsnapping van methaan ontstond een enorm broeikaseffect. Het oceaanwater werd zuurstofloos. Vijftig à zestig procent van alle tetrapoden stierf en bijna alle zeeleven. O.a. Archosaurus overleefde. Het was een vertegenwoordiger van de voorlopers van onze krokodillen, zo meent men.
Proterosuchus, een uitgestorven krokodil.
https://upload.wikimedia.org/wikipedia/commons/thumb/c/c9/Proterosuchus_BW.jpg/266px-Proterosuchus_BW.jpg
In het Boven-Perm kwamen in Zuid-Afrika en Zambia gemeenschappen voor van zoogdierachtige reptielen. Hadden ze een harige vacht? En konden ze hun lichaamstemperatuur regelen?
3.1.7 Overzicht van de eerste periode.
Als onze aarde begonnen is als Pangea vonden de orogenese-processen plaats toen de aardgebieden uit elkaar begonnen te schuiven. Tezamen met vulkaanuitbarstingen, vloedgolven en aardbevingen.
De continenten gingen uiteen. Noord- en Zuid-China werden op elkaar geperst door het opdringende India en het ontstaan van de Himalaya. Daar eveneens enorme steenkoolvelden.
Gebieden op aarde met steenkool.
http://i1.ytimg.com/vi/jULKOqylN_k/maxresdefault.jpg
Let er op: de grote steenkoolgebieden langs en in de omgeving van meestal hoge gebergten.
De opheffing van de Ardennen tijdens de Hercynische orogenese blijkt voldoende te zijn geweest voor de vorming van steenkoollagen in Nederland en in België.
De totale reserves aan steenkool bedraagt 1035 miljard ton. Daarmee kunnen we bij de huidige leveranties nog 130 jaar vooruit!
Daarnaast moet er in samenhang (??) met steenkool nog aardolie en aardgas ontstaan.
In hoeveelheden die welhaast onmeetbaar zijn. Nu nog een reserve van ong. 1.2 biljoen m3 aan aardgas!
Aardolie en aardgas moeten, i.t.t. steenkool, zijn ontstaan in de diepzee (1 à 3 km diep) (??)
Plantaardig materiaal (vooral algen) zakten naar beneden en werden door anaerobe bacteriën en aaseters omgezet in kerogeen. Dit bevindt zich b.v. in teerzanden. Als het kerogeen overdekt wordt door dikke lagen sediment kan de temperatuur oplopen tot 100 graden C. Dan ontstaat er aardolie en bij nog hogere temp. (hoger dan 120 graden C) aardgas. Een ondoordringbare laag moet tenslotte beide delfstoffen gevangen houden. Dit kan een zoutlaag zijn die ontstaan is in het Perm. Of leisteen.
Echter: tijdens het Carboon bestonden de enorme zoutlagen nog niet (?).
In het moedergesteente ontstaat de olie of het gas.
https://upload.wikimedia.org/wikipedia/commons/thumb/f/f6/Prospeccion_petroleo-nl.svg/260px-Prospeccion_petroleo-nl.svg.png
Is dit een geloofwaardige veronderstelling?
Verspreid over de hele wereld, in de loop van miljoenen jaren, deze regelmatige opbouw en omzettingen? Want ook dalingsgebieden van (on)diepe zeeën zijn voorwaarde voor ontstaan
van olie en gas. De lagen met organische afzettingen moeten dalen onder hoge druk tot 1 à 1.5 km. om aardolie te laten ontstaan. Aardolievelden vind je eveneens veelal in de streken met gebergtevorming.
Gebieden op aarde met aardoliewinning.
https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEjM7kWp0jL0A3EjmI-vveQRqG9Fc7jeTQTa0fUShkBMkYZ-KDqBR0cf2NFr5IvnqIBiS8LbjX06QHaoBxoo8QJgEjA-hf7gzYDQC0orAXn8YlxmK4pNJd58bo-dJ-Wg0TCyV3ugbVloMIhJ/s400/globalmap-oil.jpg
Wereldwijd worden dikke zoutlagen gevonden. Vaak in samenhang met olie, gas en steenkool.
De zoutlagen zijn duizenden vierkante km's groot en tot 1 km. hoog.
Bovendien zijn er zoutpilaren van 2 à 3 km hoog.
Gedurende het Perm zouden de zoutlagen moeten zijn gevormd.
De zoutformaties onder Nederland behoren tot Zechsteinformatie. Deze is 500.000 km2 groot en 1 km. dik. Ze bevindt zich onder Noor-Nederland en loopt door onder Denemarken, Noord-Duitsland, Polen tot in Rusland. Ze bevat geen vervuiling en geen fossielen!!
Onder het Great Barrièrerif bij Australië is de zoutlaag 5 km. dik en onder de Dode Zee 10 km.
De bovenste zoutlagen bestaan uit 99% NaCl en in diepere gedeelten bevindt zich CaSO4 .
Dit laatste kan niet in zeewater ontstaan. In zeewater ontstaat CaSO4.2H2O (gips).
Een prachtige creatieve weergave van zoutpilaren onder de grond.
EeEen schematische weergave van de zoutpilaren.
Volgens de gangbare theorie is het zout neergeslagen in ondiepe zeearmen. Doordat het kouder werd in het Perm (onze gebieden schoven op naar het noorden) daalde de zeespiegel.
Het water werd zodanig verwarmd dat het zoutgehalte steeg van 3.5 tot 35 %. Dan raakt het water verzadigd en slaat het zout neer op de bodem. Zou dit proces zich nu nog ooit ergens op aarde voltrekken? B.v. rond de Dode Zee?
Hier moet ook weer sprake zijn van een geleidelijke bodemdaling. En er zouden strandwallen moeten ontstaan met niet te brede en niet te smalle doorgangen. Niet te breed: dan ontstaat er gewoon eb- en vloedbeweging.
Vervolgens moet het zout naar 2 km. diep zakken. Dan wordt het plastisch en kan het binnendringen in scheuren van het bovenliggend gesteente. Terwijl intussen de daling doorgaat ontstaan de enorme zoutkoepels.
Nog wel een probleempje: de dichtheid van het zout is groter dan die van het bovenliggende gesteente.
(gegevens deels ontleend aan Ir. Stef Heerema: Vulkanische oorsprong van zoutlagen; Almere 2012.
De foto's zijn genomen uit de presentatie door Ir. Heerema gegeven tijdens die bijeenkomst).
Volgens Stef Heerema kunnen deze formaties ontstaan zijn door een machtige stoomkolom tijdens vulkaanuitbarstingen. In een zoetwaterafzetting. Hij plaatst deze gebeurtenissen in de periode rond de
Zondvloed (ong. 2300 v. Chr.).
3.2 De tweede periode (Trias, Jura, Krijt).
Dit is eveneens een duidelijk afgegrensde periode omdat er veel bewijs voor is dat aan het einde van het Krijt de tweede gigantische extinctie heeft plaats gevonden. Daarbij kwam er een einde aan het dino-tijdperk.
Zowel lopende, zwemmende als vliegende dino's verdwenen van de aarde. Niet alleen zij, ook de vele Ammonieten, Belemnieten, Rudisten (2-kleppige schelpdieren op de riffen), Foraminiferen en Benettitales.
Ong. 75 à 90 % van alle land- en zeedieren stierf uit aan het einde van deze periode.
Als oorzaak van deze massa-extinctie wordt tegenwoordig door de meeste geologen gedacht aan het neerkomen van een gigantische meteoriet in de Golf van Mexico. De sporen van die inslag zijn nog steeds aanwezig in de vorm van een grote krater onder land en zee van Mexico.
Tevens zijn in Siberië oppervlakten van 7 miljoen km2 stromen basaltlava gevonden. Ontstaan in diezelfde periode? Of al tijdens het Perm? Door ontsnapping van methaan uit de zeebodem werd het broeikaseffect nog flink verhoogd. Het zuurstofgehalte van het zeewater werd drastisch verlaagd; zo is de gedachte.
3.2.1 Trias.
In het Trias schijnt de gemiddelde temperatuur op aarde heel hoog geweest te zijn: 220 C.
Er zijn geen ijskappen. Dit zou welhaast overeen kunnen komen met de situatie van de aarde kort na de schepping.
De fossiele vindplaatsen van het vroeg-Trias blijken niet erg wezenlijk te verschillen van die van de eerste periode: afzettingen in modder in semi-aride gebieden. Tijdens het Trias keren vele ondiepe continentale zeeën weer terug! Dit door het begin van gedeeltelijk uiteenvallen van Pangea. Er zijn veel riffen rond de Tethyszee.
In Noord-Europa zien we mariene transgressies in een gebied van woestijnen, bergen en meren.
Rond de evenaar kwamen grote woestijngebieden voor en grote bekkens met veel verdamping van water.
De hele aarde vertoont dus steeds meer variatie in leefgebieden.
Veel voorkomende afzettingen waren rode zandgesteenten.
Er waren gebieden met seizoensregens in drogere gebieden van het huidige Rusland; het bekken van Moskou. Er leefden (long)vissen, amfibieën en reptielen (sauriërs).
Eenzelfde soort leefgemeenschap bestond in Antarctica. Een reusachtig groot gebied met Voltziopsis-bossen (fossiele coniferenbossen) bestaande uit vele bomen. maar zonder vorming van steenkool. Vandaar de veronderstelde wereldwijde 'steenkoolkloof'' gedurende ong. 20 miljoen miljoen jaren.
In Duitsland vond men fossielen in een kustdelta met opdrogende meren waarin de bewoners stierven. De wateren krioelden van vissen, garnalen, kreeften, trechterspinnen en degenkrabben (Limulus, nu levend in Z.O. Azië en Noord-Amerika). Degenkrabben of Molukkenkreeften zijn zgn. 'levende fossielen' die 360 miljoen jaren lang niet veranderd zijn.
In de vijvermodder werden duizenden insecten, behorend tot 200 soorten en vele arthropoden gevonden. Samen met Votzia, paardenstaarten en varens. En de "eerste" trechterspin.
Gedurende het Perm waren de Therapsiden al aanwezig. Deze dieren worden beschouwd als zoogdierachtige reptielen.
Ze hadden hun poten rechter onder het lichaam geplaatst, hun gebit was meer gedifferentieerd, de kaakspieren waren groter en de dieren konden tegelijkertijd eten en ademhalen. Dit laatste werd mogelijk door de vorming van een tweede gehemelte.
Sommige Therapsiden bezaten misschien een vacht en waren dan warmbloedig.
Het vervolg laat zich voorspellen! Gedurende Jura, Krijt en Tertiair gaan de zoogdieren steeds meer gebied "veroveren".
Er ontstaan duizenden nieuwe soorten.
In de zuidelijke Alpen op de grens tussen Zwitserland en Italië is een vindplaats met honderden complete skeletten van zeereptielen (geen kleine jongens; Ichtyosauriers, Notosauriërs en Placodonten) samen met duizenden vissen (Actinopterygii) en inktvissen. Dan nog de merkwaardig uitziende Tanystropheus.
https://upload.wikimedia.org/wikipedia/commons/thumb/4/4a/Tanystropheus_NT_small.jpg/220px-Tanystropheus_NT_small.jpg
Aan de voet van gebergten in Kirgizië heb je ook vindplaatsen van beboste overstromingsgebieden van meren en moerassen. Met duizenden (soms zeer grote) insecten (Titanoptera).
De Triasflora bestaat uit Palmvarens, Ginkgo's, Coniferen en Benettitales.
Discontinue steenkoollagen bevinden zich tussen de modderafzettingen met al die fossielen.
An de voet van de Andes in het uiterste zuiden van Zuid-Amerika bevind zich een fossielenbed van zand en modder. Nu is het een vulkanisch actief gebied van rivieren en meren; met een 40 m. dikke laag van reusachtige coniferen, varens en paardenstaarten.
Vrij beroemde dinosauromorfen (op dino's gelijkend) zijn Herrerosaurus en Eoraptor.
Pisanosaurus werd hier ook al gevonden. Dit is een zgn. Ornithischier. Van deze sauriërs stammen de vogels af; aldus meenden de geleerden in vroeger tijden. Dit moet zo zijn omdat de bekkenbeenderen van deze groep overeenkomsten vertonen met die van de vogels. De Saurischiërs hebben een totaal andere stand van hun schaambeen die overeenkomt met die van de zoogdieren.
Tegenwoordig meent men dat vogels afstammen van Saurischiërs.
Woestijnen hebben in de triastijd bestaan in Schotland. Fossielen zijn hier bedekt met opgewaaid duinzand. Hier moesten een aantal vrij grote reptielen aan voedsel zien te komen.
Er wordt gedacht dat de woestijn hun voortplantingsgebied was en het voedsel verderop werd gevonden. (Zo is voor alles een oplossing!)
Dichtbij de Rocky Mountains in Amerika waren rijk beboste overstromingsgebieden aanwezig. Met veel zand en slik. Wel 1300 gewervelden werden er gevonden. Allerlei typen Archosauriërs werden er opgegraven. Daaruit kwamen dus de latere sauriërs voort.
In de Ghost Ranch van New Mexico kwamen honderden skeletten van de beroemde Coelophysis tevoorschijn. Snelle jongens die graag vlees verorberden. Terwijl er geen planteneters aanwezig zijn waar deze roofdieren van zouden moeten leven.
De merkwaardige veronderstelling wordt gedaan dat deze door elkaar gesmeten dieren stierven door gebrek aan water en daarna door de rivier werden meegesleurd.
Coelophysis.
https://upload.wikimedia.org/wikipedia/commons/thumb/4/4a/Tanystropheus_NT_small.jpg/220px-Tanystropheus_NT_small.jpg
Opvallend: De gemeenschappen van sauriërs floreerden vooral in de laaglandgebieden, langs riviermondingen en moerassen. Daar groeiden vooral varens, wolfsklauwen en paardenstaarten.
Meest bijzonder in het boven-Trias waren de eerste schildpadden. Niemand weet waar ze vandaan kwamen. Ze ontstonden natuurlijk uit "stamreptielen". Rugschild (carapax) en buikschild (plastron) omgeven het dier. Het rugschild is samengesteld uit verbrede, vergroeide ribben.
De ledematen en de beide gordels kunnen zich binnen dit schild terugtrekken.
Er leven ong. 350 soorten die voorkomen rond de hele aardbol, uitgezonderd de noordelijke streken.
In Nederland leefde vroeger de moerasschildpad.
3.2.2 Jura.
Ook toenmaals grote ondiepe zeeën en grote delta's, lagunen en open shelfzeeën.
Als eerste een vindplaats in het uiterste zuiden van Engeland.
Fossielen achtergelaten in zuurstofarme modder op de zeebodem langs de kust; samen met kalkriffen.
Fossielen: Ichtyosauriërs, Plesiosauriërs, een Pteriosauriër, 50 soorten vissen en talloze ammonieten.
Vervolgens gaan we naar zuid-Duitsland, op de grens met de Alpen. Daar zijn meer dan 500 vrijwel complete skeletten van Ichtyosauriërs, samen met inktvissen gevonden. Nu in bitumineuze kalkstenen en schalies.
Daarenboven nog eens 70 Teleosauriërs (krokodillen).
Deze allen afgezet in enkele diepe zeebekkens (met zuurstofarm water?) van de overigens ondiepe zee die toen heel Noord-Europa bedekte.
Nu naar een afzetting in Daohugou (Binnen-Mongolië): moddergesteenten in meren- en rivierengebied met veel overstromingen in dit bergachtige land.
Hier leefde het zwemmend zoogdier Castorocauda, het "eerste" zoogdier als fossiel. (Hoe zit dat nu met de terugkeer van grote landzoogdieren naar het water?? Daardoor ontstonden immers de zeehonden, dolfijnen en walvissen!) Castorocauda had een 'beverstaart' en goede zwempoten met zwemvliezen. Daarom moet de tijd sinds het verschijnen van 'waterzoogdieren' met 100 miljoen jr. worden vervroegd!!
https://upload.wikimedia.org/wikipedia/commons/thumb/0/0a/Castorocauda_BW.jpg/266px-Castorocauda_BW.jpg
Er blijkt een compleet nieuwe familie van zoogdieren te zijn ontstaan.
Het zoogdier Volaticotherium kon "vliegen" als een vliegende eekhoorn. Perfect gebouwd met een vlieghuid tussen de vier poten.
https://upload.wikimedia.org/wikipedia/commons/thumb/5/5e/Volaticotherium.jpg/266px-Volaticotherium.jpg
Dan nog Pedopennae, een eerste gevederde sauriër (??) Er werden "vogelpoot" beenderen gevonden met grotere en kleinere veren. Er is gesuggereerd dat het dier 4 paar vleugels had: 2 om mee te vliegen en 2 aan de poten om sneller mee te rennen (?).
En dan niet te vergeten Pseudotribos, een lid van de Monotremata (buideldieren).
Daarna vinden we in het gebied van de West-Alpen (Fr.) weer afzettingen in fijne modder van een zuurstofarme zeebodem. Hier is de "eerste" octopus gevonden. En wel 3000 slangsterren per m2 .
Verder vissen, kreeften en inktvissen.
Dan gaan we het land op. Aan de oostkust van Afrika, in Tanzania, werden in riviervlakten de skeletten van reuzen-dino's ontdekt. Er werd ruim 200 ton aan beenderen naar Duitsland verscheept. B.v. Brachiosaurus; 12 m. hoog en 23 m. lang.
Hun grote poten staan rechtop in de modder! Volgens geleerden waren ze weggezakt in moerassen toen ze op zoek waren naar voedsel. Een echte sauropode.
Diverse typen dino's kwamen hier voor. O.a. ook de beroemde Rhamphorynchus, een Ichtyosauriër.
Verder de fossielen van zoogdieren, varens en coniferen.
Nu gaan we naar de kust van Portugal. Daar lagen bosmoerassen. Nu afzettingen van schalies, kalksteen en steenkool. Tienduizenden fossielen zijn hier opgegraven. Zowel van land- als water dieren.
Schelpdieren, insecten, vissen, amfibieën, schildpadden, pterosauriërs, dinosauriërs, vogels, zoogdieren (26 soorten) en een zeekrokodil. Het hele dierenrijk is bijna vertegenwoordigd!
Daarbij nog tanden van Archaeopteryx.
In het zuiden van Duitsland, aan de rand van de Alpen, in het beroemde Solnhoven, werd een echte veer gevonden. Dus vanaf 150 miljoen jr. geleden, zeggen de geologen, waren er vogels op aarde.
Later werd hier ook Archaeopteryx ontdekt. Eindelijk sluitend bewijs dat vogels van dino's afstammen? Is er een eerste echte 'missing link' gevonden? Dat valt nog te bezien!
Er zijn intussen 10 exemplaren van Archaeopteryx ontdekt. Ze hielden niet van leven in bomen omdat ze geen opponeerbare eerste teen hadden, zoals alle vogels hebben.
Duizenden fossielen zijn in Solnhofen gevonden in de zuurstofarme modder van de kustlagunen. Tijdens de moessonregens en cyclonen verdronken vele grote landdieren.
Maar ook grote Pterodactyli (Rhamphorynchus). Compsognatuss, Inktvissen, samen met kreeften (garnalen), insecten en vogels kwamen om.
Midden in de Rocky Mountains ligt een gebied van 1.5 miljoen km2 : de Morrisonformatie. Het was een gebied van woestijnen, meren, rivieren en moerassen. De fossielen zitten nu in zand en modder. Supergrote dino's kwamen om. Verklaring: de ondiepe zee trok zich terug en duizenden dieren verdrongen elkaar in verlangen naar water. Nu lagen er o.a. 44 exemplaren van Camarosaurus, Diplodocus, Apatosaurus, Camptosaurus en Stegosaurus, Zoek ze allemaal maar op.
Diplodocus spec.
https://upload.wikimedia.org/wikipedia/commons/thumb/4/44/SeismosaurusDB.jpg/220px-SeismosaurusDB.jpg
In een nabijgelegen gebied kwamen ook Supersaurus en Ultrasaurus voor. Daarenboven: pterosaurirs, waterkrokodillen, schildpadden, amfibieën, longvissen en allerlei soorten zoogdieren !
Samen met de ons vertrouwde planten: mossen, levermossen, paardenstaarten, varens, palmvarens, cycas, ginkgo's en coniferen (waaronder Araucarites. De huidige slangenden behoort tot deze fam.).
Araucaria of apenverdriet.
https://upload.wikimedia.org/wikipedia/commons/thumb/3/38/Araucaria_araucana1.jpg/266px-Araucaria_araucana1.jpg
3.2.3 Krijt.
Hiermee maken we de overgang naar het Krijt.
Een wel heel bijzondere periode waarbinnen bijzondere ontwikkelingen zouden plaats vinden van zoogdieren, vogels en angiospermen (bedektzadigen).
In Engeland en Frankrijk werden de skeletten van fuutachtige vogels en van gansachtige vogels opgediept.
In het onder-Krijt zijn de geslachten van populieren, vijgen en laurierbomen al vertegenwoordigd.
Dit betekent dat ontwikkeling van deze groepen al eerder, in het Carboon, moet hebben plaats gevonden.
Er is één paleobotanicus (wie??) die geschreven heeft: het ontbreken van fossielen van deze geslachten komt misschien voort uit het feit dat angiospermen zich in hoger gelegen gebieden hebben ontwikkeld, waar conservering niet erg waarschijnlijk is!
We beginnen dicht bij huis; de kust van België. Hier werden de 30 complete skeletten gevonden van Iguanodon, die nu in Brussel te bewonderen zijn. Ze kwamen terecht in de modder van een zoetwatermeer. Werden ze door een overstroming het meer ingesleurd en bleven ze daar bewaard dankzij de koude bodemafzettingen? Een ongeloofwaardig verhaal!
https://upload.wikimedia.org/wikipedia/commons/thumb/2/26/Iguanodon_28-12-2007_14-18-33.jpg/260px-Iguanodon_28-12-2007_14-18-33.jpg
Verder weg in een noordelijke vlakte van China bij Liaoning, omringd met vele vulkanen, werden prachtige fossielen opgediept. Weer uit de modder van meren. Bedolven onder regens van as!
De rijke voorraad bestond uit Pterosauriers, zoogdieren en vogels.
Uitvoerig bediscussieerd werden de "veren" van Sinosauropteryx. Waren de "veren" slechts collageen-uitsteeksels op de huid?
Een beroemd zoogdier uit die tijd is Repenomanus. Een compleet skelet werd gevonden.
Fossielen van zoogdieren in Jura en Krijt zijn een uitzonderlijke zeldzaamheid!

https://upload.wikimedia.org/wikipedia/commons/thumb/b/bd/Repenomamus_BW.jpg/266px-Repenomamus_BW.jpg
Het eerste buideldier dat fossiel bewaard werd was Eomaia.
Er leefden echter ook diverse soorten sauriërs die veren hadden. Veren aan de "handen" en/of aan de poten. Een beroemd v.b. is de Microraptor. Honderden complete skeletten van Microraptor werden gevonden, perfect bewaard onder de vulkaanas. Wikipedia wijdt een heel uitvoerige beschrijving aan deze dieren en hun verwanten.
https://upload.wikimedia.org/wikipedia/commons/thumb/4/44/Microraptor_by_durbed.jpg/260px-Microraptor_by_durbed.jpg
De verschillende soorten van het geslacht Microraptor worden gerekend tot de fam. Dromaeosauridae. Vertegenwoordigers hiervan werden gevonden verspreid over de hele wereld! Ze hadden veelal wel echte veren maar konden niet vliegen als vogels. De veren zouden hebben gediend als pronkstukken bij paargedrag, het sneller kunnen rennen of het vasthouden van warmte bij de warmbloedige sauriërs.
Een beeld van een Dromaeosauride.
https://upload.wikimedia.org/wikipedia/commons/thumb/7/7c/Utahraptor_BW.jpg/266px-Utahraptor_BW.jpg
Deze dieren leefden in een weelderige begroeiing van de ons intussen welbekende planten. Maar daarbij kwamen ook de eerste bloeiende planten, angiospermen dus voor.
Door Chinese boeren werden duizenden fossiele "vogels" verzameld. Ze kregen de naam Confuciusornis. Ze worden zeer uitvoerig besproken in Wikipedia.
Ze konden waarschijnlijk niet vliegen. Ze hadden geen kiel op hun borstbeen voor de aanhechting van grote vliegspieren. Het uiteinde van de arm bevatte klauwtjes. Ook waren de 'vleugelbeenderen' minder met elkaar vergroeid dan bij moderne vogels. Waren het loopvogels die in bomen konden klimmen ?
https://commons.wikimedia.org/wiki/File:
Confuciusornis_sanctus_(2).jpg
Heel bijzonder: er zijn van deze "vogels" meer fossielen gevonden dan van alle andere dino-soorten samen. Terwijl ze toch prima zouden moeten kunnen ontsnappen tijdens natuurrampen!
Binnen een oppervlakte van 10 m2 werden 40 "vogels" gevonden; dicht naast elkaar.
https://upload.wikimedia.org/wikipedia/commons/thumb/b/be/Confuciusornis_plumage_pattern.jpg/260px-Confuciusornis_plumage_pattern.jpg
In een vergelijkbaar gebied, niet ver daar vandaan, werden duizenden botten gevonden. O.a. 500 kleine zoogdierfossielen, verdeeld over 17 soorten. Van 'primitief' tot geavanceerd zoals Prokenalestes, een buideldier.
In een zoetwaterlagune aan de kust van Brazilië werden honderden nieuwe soorten ontdekt in de fossiele modder. En naast dichotoom vertakte sporenplanten vertegenwoordigers van de plantenorde Nymphaeales waartoe o.a. de waterleliefamilie behoort.. Ook andere angiospermen kwamen voor; samen met vele insecten voor de bestuiving.
Samen met spinnen, insecten, schildpadden, hagedissen, Pterosauriers, vogels en vissen.
Prachtig versierde helmen van grote Pterosauriers.
https://upload.wikimedia.org/wikipedia/commons/thumb/5/56/Tapejara_heads_BW.jpg/350px-Tapejara_heads_BW.jpg
In de zgn. Dinosaur Cove. aan de kust dicht bij Melbourne (Australië) lagen duizenden botten van kleine ornithopode dino's. Een groep sauriërs als hieronder afgebeeld. Tegenwoordig spreekt men liever van Euornithopoda; dit betekent 'echte lopers op 2 poten zoals de vogels'.
https://upload.wikimedia.org/wikipedia/commons/thumb/b/be/Ornithopods_jconway.jpg/260px-Ornithopods_jconway.jpg
Australië was in die tijden nog verbonden met Antarctica en dus met het zuidelijke Gondwanaland. Maar het klimaat op Antarctica was nog gematigd (120 C) en er was voldoende regenval voor uitbundige plantengroei. Ook zeereptielen zoals Kronosaurus (13 m. lang) liggen hier begraven.
https://en.wikipedia.org/wiki/File:
Kronosaurus.jpg
Eén van de grote problemen binnen de geologie blijft hoe je echte vogels moet laten ontstaan uit sauriërs. Alleen de stand van het schaambeen (pubis) in de bekkengordel kan toch niet een doorslaggevend argument zijn om de vogels te laten ontstaan uit de zeer heterogene groep van Ornithischiers. Zoals vroeger gedaan werd. Omdat de stand van het schaambeen bij vogels vergelijkbaar is met die van Ornithischiers.
https://upload.wikimedia.org/wikipedia/commons/thumb/e/eb/Ornithischia.png/266px-Ornithischia.png
Dit in tegenstelling tot de Saurischiers, waarbij het schaambeen naar voren wijst.
https://upload.wikimedia.org/wikipedia/commons/thumb/8/8d/Saurischia.png/266px-Saurischia.png
Onze huismus zou dan tot de Therapoda behoren. Onze vogels worden dan Neornithes genoemd.
De Therapoda en dus ook Neornithes behoren tot de Saurischiers.
Grappig is wel om te zien hoe de huismus totaal vereenzaamd tussen zijn/haar verwanten voorkomt.
Als je een groot 'evolutie-geloof' bezit kun je het misschien wel plaatsen.
https://upload.wikimedia.org/wikipedia/commons/thumb/4/45/Various_dinosaurs3.png/266px-Various_dinosaurs3.png
De hele wereld van Sauriërs komt voort uit het Trias. Let er op dat de het ontstaan van de vogels en verwanten erg onzeker is.
Wil je bijna alles weten over theorieën over de evolutie van vogels kijk dan op
'oorsprong van de vogels ' bij Wikipedia.
In een gebied in het uiterste zuiden van Zuid-Amerika, tegen het Andesgebergte gelegen, bevond zich een deltagebied met vele rivieren en meren. In 85 m. zandsteen-, slik- en modderlagen werden alle mogelijke skeletten gevonden van o.a. Titanosauriden, Argentinosaurus, krokodilachtigen, slangen, zoogdieren etc. In deze afzettingen bevonden zich duizenden nesten van dino's met zelfs prachtig geconserveerde embryo's.
Tezelfdertijd ontstonden in Midden-China (Mongolië) afzettingen in woestijnachtige gebieden. In het duinzand en modder bleken skeletten van 200 dino's en 1000 zoogdieren verborgen. Ook Oviraptor ('eierdief'') en Velociraptor leefden hier. Waarschijnlijk stal Oviraptor geen eieren maar verdedigde z'n nest tegen een Protoceratops.
Nog zo'n vindplaats van honderden, ook zeer grote dino's, maar nu in het westen van Amerika op de grens van Canada en de VS. Gelegen aan een vroegere zeeweg, nu ten oosten van de Rocky Mountains.
Tyrannosauriden, Ceratopsiden, Hadrosauriers en Pterosauriers waren aanwezig, samen met alle mogelijke waterdieren en 20 soorten zoogdieren.
Supersaurus werd in 1972 ontdekt en was 30 m. lang en woog 100 ton!
En: veel bloeiende planten zoals eiken, lelies en zonnebloemen.
Twee afbeeldingen van Quetzalcoatlus. De grootste Pteriosaurier met een spanwijdte van 10 m.
https://upload.wikimedia.org/wikipedia/commons/thumb/7/7a/Quetzalcoatlus_by_johnson_mortimer-d9n2b06.jpg/260px-Quetzalcoatlus_by_johnson_mortimer-d9n2b06.jpg
Tenslotte komen we aan in Nederland met de zeer beroemd geworden Maashagedis, de Mosasaurus.
Napoleon vond het nodig om het skelet te stelen en naar Parijs over te brengen. Vijfentwintig geslachten van de Mosasauriërs leefden verspreid over de hele wereld.
Op een andere plek in de VS, ten oosten van de Rocky's, bevond zich eveneens een overstromingsgebied met zand, modder en veen, genaamd Hell Creek. Dit blijkt een laag die over de hele wereld is aangetroffen en die opvallend rijk is aan iridium. Deze iridiumpiek wordt in verband gebracht met de inslag van een groot object vanuit de ruimte. En moet daarmee een voorbode van de grootste extinctie van einde-Krijt zijn geweest. De weiden van palmvarens, varens en palmen zou vervangen zijn door die van honderden plantensoorten van bedektzadigen, waaronder bossen van bedektzadige bomen.
Nu blijken er wel 100 soorten kleine zoogdieren te zijn. Ze leefden blijkbaar samen met Tyrannosaurus rex!!
Hoe kwam er een einde aan het Krijt-tijdperk?
In zijn boek 'De opkomst en ondergang van de Dinosauriërs' geeft Steve Brusatte een levendige en uitvoerige beschrijving van een aarde- en hemelschokkende gebeurtenis.
Enorme lichtflitsen volgden elkaar op in Hell Creek. Het aardoppervlak deinde op en neer als een springkussen. Gloeiend hete brokken daalden neer, wouden vlogen in brand, enorme tsunami's spleten de kustgebieden. Zwavel regende neer. Jarenlange kou door zonsverduistering volgde. Dus voedselketens stortten in. Zure regens veroorzaakten nog meer rampen.
De oorzaak was een komeet of een asteroïde die neerstortte op Yucatan-schiereiland van Mexico.
Met de energie van ongeveer 12 miljard waterstofbommen. Tot 40 km. diep in de aardkorst!
Tsunami's trokken rotsen los en wierpen die over het land. In India bleven vulkanen nog 1000 jaren lang uitbarsten. Jarenlang bleef de aarde in kou en duister gehuld.
In een kleilaagje tussen Krijt- en Paleogeenafzettingen bleek een zeer hoge doses iridium aanwezig te zijn. Dit element is uiterst zeldzaam op aarde maar komt veel meer in de ruimte voor.
Kort voor de jaren negentig van de vorige eeuw werd de krater ontdekt. Een 180 km. breed gat onder dikke lagen sediment en voor een groot gedeelte onder zeewater. Gevormd tegen het einde van het Krijt.
Fossiel van een reusachtige ammoniet. 
https://upload.wikimedia.org/wikipedia/commons/thumb/0/09/Parapuzosia_seppenradensis_4.jpg/260px-Parapuzosia_seppenradensis_4.jpg
Zowel land- als zeedieren stierven grotendeels uit:
- Ammonieten (inktvissen in een spiraalvormige schelp),
- Belemnieten (tienarmige inktvissen)
- Rudisten (tweekleppige schelpdieren).
Opvallend is wel dat de grote Stegosaurussen al aan het einde van de Jura-periode uitstierven.
En dat krokodillen, schilpadden en vele boom- en plantensoorten in leven bleven.
Vrij ingrijpende milieuveranderingen door het verschuiven van de continenten kunnen een grote rol gespeeld hebben. Door verregaande regressie van de zeeën in het boven-Krijt en daardoor sterke temperatuurdaling in het tropische gebied van de Tethyszee, samengaand met sterk veranderde zeestromingen, kan het milieu van land en zee beïnvloed hebben.
Zo kunnen enorme Sequoia-wouden overgegaan zijn in gemengde loofbossen.
Daarbij komt nog de overweging dat verdergaande gebergtevorming grote invloed kan hebben gehad.
3.3 De derde periode; Kaenozoïcum.
Na de grote extinctie zouden de open gevallen niches snel zijn bezet door vogels en zoogdieren.
Een reuzenvogel, Diatryma (Gastornis), werd gevonden in Wyoming en West-Europa. Het dier was
2 m. hoog, had een kop ongeveer zo groot als een paard en geweldige looppoten met enorme klauwen.
Diatryma.
https://upload.wikimedia.org/wikipedia/commons/thumb/c/c3/Gastornis%2C_a_large_flightless_bird_from_the_Eocene_of_Wyoming.jpg/240px-Gastornis%2C_a_large_flightless_bird_from_the_Eocene_of_Wyoming.jpg
In de beroemd geworden fossiele afzettingen van de London Clay werd een rijke verzameling aan haaienskeletten gevonden. Na de uitsterving van de grote zeereptielen zoals de Ichthyosaurussen, de Plesiosaurussen en de Mosasaurussen kregen de haaien alle speelruimte in alle wereldzeeën. Verder bevatte de 'clay' alle mogelijk soorten delen van zeedieren, o.a. zeeschildpadden en krokodillen, creodonten (roofdieren), hoefdieren, primaten en het eerst "paard": Hyracotherium. Uit fossielen blijkt een grote rijkdom aan vogelleven: veel grote watervogels als pelikanen, aalscholvers en Jan van Genten.
Ook patrijzen, steltlopers, reigers, koekoeken en duiven.
(Opm. het bodemleven in de ondiepe zeeën vertoont aardige overeenkomsten met die van het Ediacaratijdperk van het Cambrium).
In deze lagen werden eveneens meer dan 300 soorten zaden en vruchten gevonden, vooral van tropische soorten.
In andere gebieden op aarde bleken tijdens het Eoceen rallen, kraanvogels en eendachtigen, gieren en (reuzen)pinguïns te leven.
In Quercy (Zuid-Frankrijk) b.v. werden zeer vele fossiele vogels opgedolven: waadvogels, ibissen, uilen, haviken, patrijzen, zwaluwen, gierzwaluwen, rallen, scharrelaars, reigers, steltlopers, scharrelaars en hoenderachtigen.
Ook in Frankrijk: hoppen, papagaaien, futen, spechten, kraaien, vinken, flamingo's, struisvogels, rhea's, tinamoes en alken.
In al weer een vergelijkbaar gebied ten oosten van de Rocky's (Crazy Mountain), in eenzelfde overstromingsgebied werden drie groepen zoogdieren aangetroffen: multituberculaten (lijkend op knaagdieren), marsupialen (buideldieren) en placentalen (dieren met placenta). Er waren al 29 families van de placentalia.
Er groeiden Plataan, Rhamnus, Vitis en Taxodium.
Uit afzettingen in het Bighorn-bekken, iets verderop, blijkt een snel herstel van angiospermen en zoogdieren. Vele typen zoogdieren komen voor, vergelijkbaar met de huidige.
Grote temperatuurschommelingen kwamen voor; variërend van 4 tot 26 graden C als jaargemiddelde.
Toen de Rocky Mountains werden gevormd ontstonden er enorme oppervlaktes met meer-sedimenten (65.000 km2 ) Ze waren wel 2 km. dik!! In de afzetting van Green River werden minstens 1 miljoen fossielen verzameld. Alle klassen van schimmels t/m zoogdieren zijn vertegenwoordigd. Paarden, tapirs, neushoorns en vleermuizen. O.a. Uintatherium, zo groot als een olifant. En vleermuizen.
https://upload.wikimedia.org/wikipedia/commons/thumb/0/0f/Uintatherium_DB.jpg/266px-Uintatherium_DB.jpg
Neogeen.
In het Neogeen (tijdperk van paleoceen t/m Oligoceen) van Zuid-Engeland heerst een tropische sfeer met regenwouden en mangrovekusten. Naast de hierboven genoemde komen Primaten (b.v. Cantius) voor en 300 soorten angiospermen.
In de carbonaatmodderlagen in Italië ten zuiden van de Alpen, vond men 500 soorten land- en zeedieren. Van de vissen 90 families, waarvan 80 nu nog bestaan, die leven in de Oceanen.
Door groei-explosies van algen zouden veel vissen gestikt zijn!(??)
In Messel (Midden-Duitsland) bestonden vergelijkbare omstandigheden rondom een meer met tropische begroeiing (dennen, beuken, kastanjes en eiken). Paarden, primaten, miereneters, knaagdieren, vleermuizen en egels trokken hier rond. Samen met salamanders, kikkers, schildpadden, leguanen, slangen en krokodillen. Rallen, ibissen, uilen en gierzwaluwen verlevigden het landschap.
De subtropische kalksteenkarst van het kustgebied in Noord-China bevatten kaakbeenderen en enkelgewrichten van antropoïden. Hieruit ontstonden. zo meent men, later de apen, mensapen en mensen.
De dageraad-aap uit China: Eosimas sinensis.
https://upload.wikimedia.org/wikipedia/commons/thumb/a/a5/EosimiasDB15.jpg/260px-EosimiasDB15.jpg
Bij Fayum, tussen de armen van de Nijl, leefden veel, ook nu nog voorkomende watervogels. Daarnaast grote "neushoorns", "olifanten" en "apen".
Dankzij vulkaanuitbarstingen bleven prachtige bloemen en vlinders bewaard rond een groot meer in het midden van de VS (Florissant). Wel 1500 soorten insecten zorgden voor de bestuiving.
Vogels en zoogdieren waren er heel zeldzaam, terwijl amfibieën en reptielen er niet voorkwamen.
Op de hogere gebieden: pijnbomen en eiken.
Als we nu naar Centraal Azië (Mongolië) gaan komen we in een gebied van grote grasvlakten. Via het Eoceen zijn we in het Oligoceen beland (30 miljoen jr. geleden). Hier leven geen "neushoorns", "tapirs" en "beren". Wel "varkens", "hyaena's", "herten", herkauwers, roofdieren, katten en civetkatten.
Het grootste landzoogdier werd hier gevonden: Indricotherium. Schouderhoogte 5.5 m., lengte 8.5 m.
https://upload.wikimedia.org/wikipedia/commons/thumb/3/30/Indricotherium.png/266px-Indricotherium.png
Het eerstvolgende gebied van het Neogeen (23 miljoen jr. geleden?) ligt in het kustgebied van Noord-Australië. Nu lagen de fossielen in carbonaat modder op de bodem van een meer.
Meren en rivieren vol met schildpadden, kikkers, palingen, longvissen etc. En krokodillen. De buideldieren domineerden: opossums, kangoeroes, Priscileo (een buidel-"leeuw"), wallaby's en buidelmollen.
Neohelos (een buideldier ?) was een nijlpaardachtige planteneter die op geen enkel ander dier lijkt.
Vergelijkbare overstromingsgebieden waren er in Midden-Afrika, nu gecombineerd met vulkanische as. Daardoor veel complete skeletten van runderachtigen, knaagdieren, lori's en mensapen (Proconsul) enz.
Ten zuiden van de Himalaya bevinden zich 2-5 km dikke sedimentlagen met duizenden aggregaten van losgeraakte botten. Met alle soorten waterdieren, landzoogdieren (50 of meer soorten "nijlpaarden", "herten", "hyena's") en vogels.
Vroege "olifanten" en mensapen (Sivapithecus) liepen er rond of klommen in de bomen.
Rond een groot meer in Centraal Afrika leefde de hominide Sahelanthropus. Zij leefden er te midden van grote "tijgers", "elanden" en everzwijnen. De woestijn bestond hier, nog samengaand met een strook bos, rond een meer.
Rond de Riftvallei in Afrika bestond een savannegebied met bossen. Bedekt met vulkanische as. Hier bleven menselijke voetafdrukken bewaard en slechts een kaakbeen van Australopithecus afarensis. Verwanten werden opgediept in Ethiopië: de Lucy-mensengroep.
Kwartair.
Zuid-Afrika (Sterkfontein): savanne en bos. Fossielen in kalksteengrotten. Waaronder 500 hominiden (Australopithecus africanus) en vele grote zoogdieren. 2.8 miljoen jr. oud.
Afrika; de Alduvaikloof: savanne met meren en rivieren. Paranthropus boisei (een planteneter) en Homo habilis (een omnivoor), samen met afgeronde stenen die dienden als gereedschap? 1.8 milj. jr. oud.
Noord-Spanje: karstkalksteen. Homo antecessor of Homo erectus of Homo heidelbergensis. Ook nu weer met steenbrokken die dienden als gereedschap? 780.000 jr. oud.
China: kalksteengrot. Daarin schedels van 40 hominiden: Sinanthropus pekinensis. Samen met makaken. En: primitieve gereedschappen (steenfragmenten). 500.000 jr. oud.
Zuid-Afrika: kalksteengrotten bij Bomblos. Homo sapiens gebruikte gereedschap en rode oker als versiering. Ze maakten ook sieraden van schelpen. 75.000 jr. oud.
Zuid-Spanje: kalksteengrotten. Homo neanderthalensis. Ze bewoonden heel Europa tot aan de Oeral. 50.000 jr. oud. De eerste Australiërs trokken vanuit Afrika via Zuidoost-Azië naar Australië.
Siberië: In de permafrost werden mammoeten gevonden. Jonge en oude dieren verdronken in een vijver? Samen met paarden? 40.000 jr. oud.
La Brea: een kustvlakte bij Los Angeles met teerputten. 166.000 sabeltandtijgers vonden hier de dood. Doordat vastzittende planteneters tijgers, coyotes, wolven en vele roofvogels aantrokken. Aldus het sprookjesverhaal. 1 miljoen botten werden opgediept: 58 zoogdiergeslachten (waaronder vleermuizen, muskusossen, elanden, kariboes en veelvraten)), 138 vogelsoorten, 24 reptielen. Daarnaast restanten van 80 plantentaxa. 20.000 jr. oud.
Zuid-Frankrijk/Italie: kalksteengrot met de ingang 40 m. beneden zeeniveau. In de rotswand met tekeningen van dieren. 20.000 jr. oud.
Flores: kalksteengrot waarin de Homo (?) floresiensis leefde. Deze hominide had een schedelinhoud van 400 cc, te vergelijken met die van een chimpansee. 15.000 jr. oud.
De eerste Amerikanen woonden toen in Chili aan een rivieroever met veel zand, modder en grind.
10.000 jr. geleden trokken ze naar de prairies om op buffels te jagen.
700 jr. geleden stierven de reuzegrote Moa's in Nieuw-Zeeland uit.
Ontstaan van mensen: tijdens het Eoceen kwamen de Prosimiï (halfapen) tevoorschijn. Dit waren de lemuren, lori's en spookdiertjes. Daaruit ontstonden de apen, mensapen en de mensen.
De primaten zouden, zo denken de geologen, zijn ontstaan uit kleine insecteneters.
Ergens rond de 25 miljoen jr. geleden zou dan de aftakking zijn ontstaan die leidde tot het ontstaan van de hominiden; waaronder de mensen.
De hominiden splitsten zich af van de mensapen: gibbons, orang-oetans, gorilla's en chimpansees.
Bij de mensapen kwamen de volgende evolutionaire tendensen voor, denkt men: toename lichaamsgrootte en grootte van de hersenen. Eveneens grotere hoektanden en langere en krachtiger armen. Uit deze dieren zou Ramapithecus voortgekomen zijn. Met kleinere hoek- en snijtanden en een platter gezicht. Dat gaat dus meer op een mens lijken! Wil je hem/haar zien lopen, rennen, eten, jagen etc. zoek maar een leuke film op internet.
Hoofdregel om mens te worden: breder bekken, aanpassingen aan de wervelkolom voor een opgerichte gang, gebruik van armen en handen voor technische handelingen en grotere hersenen.
De Australopithecinen "ontstonden" in Afrika als afstammelingen van Ramapithecus. Richard Moody schrijft dat ze mensaapachtig waren in schedelvorm en hersengrootte maar mensachtig in hun opgerichte gang.
Na later gevonden fossielen wordt Ramapithecus Sivapithecus genoemd. Stukken bovenkaak werden gevonden van ong. 10 milj. jr. geleden.
Uit één van deze "apen" ontstond het geslacht homo. Homo habilis ( de "knutselaar") leefde in het Quartair (ong. 1.8 milj. jr. geleden) in de Olduvai-kloof. Hij bezat uit keien bestaand gereedschap (opm.: voor alle mensapen niet ongebruikelijk!).
Ietwat later is, denkt men, Homo erectus ontstaan in. Zijn skelet leek wel op dat van ons maar de hersenomvang was 25% kleiner. Hij had zware wenkbrauwen maar geen voorhoofd.
Homo erectus. https://upload.wikimedia.org/wikipedia/commons/thumb/d/de/Sangiran_17-01.JPG/266px-Sangiran_17-01.JPG
Vanuit Afrika verspreidde deze soort zich over een groot gedeelte van Azië tot in Indonesië, alwaar Eugene Dubois het eerste "exemplaar" vond op Java; de 'javamens'.
De 'Pekingmens' (Sinanthropus pekinensis) leefde eeuwenlang in grotten. Ze onderhielden vuren. In de grotten bleven grote hoeveelheden botten van herten, olifanten, neushoorns en mensen bewaard. Was dit laatste een gevolg van kannibalisme?
(Opm.: zou het ook kunnen zijn dat Homo sapiens dit resultaat opleverde en zelf met de 'noorderzon' is vertrokken?).
In Syrië en in Frankrijk werden vergelijkbare grotvondsten gedaan.
Dankzij vulkanische as weten we dat er in de omgeving van de grote meren in Afrika (het Rifgebied) aapachtige wezens rondliepen. Man en vrouw? Hun voerafdrukken werden gevonden. Ze worden wel genoemd: Australopithecus afarensis. Ook wel genoemd: Lucy's volk. Omdat een beroemd geworden fossiel in Ethiopië Lucy werd genoemd. Een overtuigend begin van menswording? Ze wandelden rond tussen alle grote zoogdieren van de Afrikaanse savanne. 3.5 miljoen jr. geleden (?).
 skeletonderdelen van Lucy. Ontdekt door Louis Leakey.
skeletonderdelen van Lucy. Ontdekt door Louis Leakey.https://upload.wikimedia.org/wikipedia/commons/thumb/0/0e/Lucy_Mexico.jpg/266px-Lucy_Mexico.jpg
Meer dan 500 fossielen van hominiden werden ontdekt in grotten bij Sterkfontein (Zuid-Afrika).
Uit de Olduvai-kloof, in de nabijheid van Australopithecus afarensis, werden door Leakey meer hominide skeletten gevonden. O.a. die van Homo habilis. Mooi afgeronde keistenen moeten hun menselijkheid aantonen.
Daarbij komen nog fossielen uit Spanje (Homo antecessor of Homo erectus), Engeland en China (Sinanthropus pekinensis, nu Homo erectus genoemd).
Hieronder een overzicht van vermeende menselijke afstamming:
https://upload.wikimedia.org/wikipedia/commons/9/9c/TreeS.pdf
Als toegift wat wel genoemd wordt een 'geologische openbaring': de Grand Canyon.
Bestaande uit carbonaten en zandsteen.
Er bestaan verschillende theorieën over het ontstaan van de Grand Canyon.
Dat een vrij kleine rivier als de Colorado dit enorme gesteentemassief (448 km lang, 1830 m diep en 0.5 tot 15 km breed) zou hebben uitgesleten in de loop van miljoenen jaren (vanaf 1.8 miljard jr. geleden tot nu) is wel heel onwaarschijnlijk.
Nu kunnen er in de loop van de jaren enorme watervloeden, aardbevingen, opstijging van de vlakten etc. geweest zijn, maar dan nog ......... De aardlagen tonen prachtige gelaagdheid!
In volgorde van onderaf:
precambrische metamorfen en microfossielen
cambrium
devoon
carboon
vroeg-perm
midden-perm
laat-perm
kwartair met luiaardfossielen.
Waardoor is de opeenvolging zo uiterst merkwaardig? Het hele Mesozoïcum ontbreekt! En vele andere perioden van de aardgeschiedenis!
Als het door water gebeurd is moet dit toch in elk geval gedurende vrij korte tijd in vrij zachte modder/zandlagen gebeurd zijn. En dan nog dankzij geweldige watervloeden. Komend vanaf ....... ??
https://th.bing.com/th/id/OIP.2sQgS00ebOYJPA5HfaQuKwHaFM?w=260&h=182&c=7&o=5&pid=1.7
3.4 Enkele opmerkelijke gebeurtenissen in de aardgeschiedenis.
3.4.1 Orogeneseprocessen.
1. De Caledonische orogenese. 450 - 390 milj. jr. geleden;
dus tijdens Siluur/Devoon.
Door de collisie van Baltica (Scandinavië) met
Laurentia (Noord-Amerika).
Daardoor ontstond het 'Old Red Sandstone' gebied.
De nu nog aanwezige gebergten zijn te vinden in Schotland,
Ierland, Noord-Engeland,
de VS (de Appalachen) en op New Foundland.
2. De Hercynische orogenese. 360 - 280 milj. jr. geleden;
dus tijdens Carboon/Perm.
Tijdens de vorming van Pangea.
Daarbij ontstonden de gebergten van Midden-Europa
van Spanje tot de Oeral:
het Zwarte Woud, Bohemen, de Ardennen en de Oeral.
De laatste door de botsing tussen het toenmalige
Siberië en Pangea.
3. De Jurassische orogenese. 160-145 milj. jr. geleden.
Toen was o.a. de Zuidwalvulkaan actief. Gelegen tussen Harlingen en
Vlieland. Ze is nu bedekt met 2000 m. sediment.
Het Zuidwalgasveld levert nu altijd nog 6.5 miljoen m3 aardgas per dag!
De gemiddelde temp. is op 2000 m. diepte 135o C i.p.v. 100o C.
In die tijd bewogen Turkije en Iran Noordwaarts.
Als gevolg daarvan vulkanisme in Europa.
Dankzij de afdekking van een vulkaan met zandsteen kan het gas worden opgevangen in de holtes en duizenden jaren lang bewaard. Het gas wordt vastgehouden dankzij een afsluitende laag kleisteen.
Een zeer wonderlijke samenloop van processen die snel verlopen moeten zijn!
Het is een vrij onzinnige gedachte dat een dergelijk systeem tot stand zou komen door geleidelijke processen van miljoenen jaren met een regelmatige opeenvolging van ophoping van organisch materiaal en bedekking met vrij kleine lagen sediment.
https://i.pinimg.com/originals/e5/79/02/e579024161b16f1ce5d22e80ccd2b5f5.jpg
Een tweede vulkaan, actief in dezelfde periode, is ontdekt in het Nederlandse deel van de Noordzee. Gelegen 100 km. ten Noordwesten van Terschelling. Hier is de vulkaan bedekt met 3300 m. sediment.
De activiteit van de vulkaan ging gepaard met zoutvloei. Dit lijkt in overeenstemming met de opvatting van Ir. Stef Heerema als het gaat over de vulkanische oorsprong van de enorme zoutlagen onder N.W.-Europa.
https://images1.persgroep.net/rcs/EHSrXddR6ToLw9WsTJPDoHEpg_o/diocontent/170445776/_fitwidth/763?appId=93a17a8fd81db0de025c8abd1cca1279&quality=0.8
4. De Alpiene orogenese. 65 milj. jr. geleden; aan het einde van het Krijt.
Afrika en India schuiven noordwaarts en
Noord- en Zuid-Amerika westwaarts.
Zo ontstaan: de Pyreneeën, het Zagrosgebergte,
de Rocky Mountains, de Alpen en de Andes.
In het studieboek Earth History and Palaeogeography (door H.Torsvik, L.Robin en M.Cocks) worden 48 verschillende grote en kleine orogenese-processen beschreven. Al die gebeurtenissen samen kunnen het voorkomen de vele fossielen-kerkhoven wellicht verklaren!
3.4.2 Perioden van massa-extincties.
Het frappante is dat er eveneens vier perioden van massaextinctie worden beschreven.
Er leven nu nog ong. 1% van alle soorten organismen die ooit op aarde hebben geleefd. De oorzaak daarvan wordt vooral gezocht in de CO2 - cyclus. Dit vooral door de zgn. LIP-erupties (Large Igneous Province). Tijdens perioden met enorme vulkanische activiteit ontstonden vulkanische scheuren die er voor zorgden dat 100.000-den km2 werden bedekt met gesmolten gesteenten. Daarbij kwamen geweldige hoeveelheden CO2 vrij die zorgden voor temperatuurverhoging en verzuring van de oceanen. De oceanen werden daardoor anoxisch en in zee levende organismen kwamen grotendeels om. Na een overweldigende algenbloei, die giftig was voor de zeedieren en na afsterving alle O2 verbruikten tijdens de afbraak.
Allemaal processen die in betrekkelijk korte tijd verliepen!
1. Extinctie tijdens Ordovicium-Siluur. 444 milj. jr. geleden.
Vgl. met de Caledonische orogenese.
IJs overdekte toen de Zuidpool en de zeespiegel daalde met 150 m.
De daaropvolgende zeespiegelstijging veroorzaakte anoxie
en er kwamen giftige metalen vrij uit de sedimenten; denkt men.
85% van de organismen kwam om. Vooral: koralen, Brachiopoden,
Trilobieten en Conodonten. Van de Conodonten worden grote
hoeveelheden zoogdierachtige tanden/kiezen gevonden.
2. Extinctie tijdens het Laat-Devoon. 380-360 milj. jr. geleden.
Vgl. met de Hercynische orogenese.
75% van de organismen kwam om. Vooral Goniatites (inktvissen)
en zeesponzen. Opnieuw door anoxie.
In de tijd ontstonden de Viljoentrappen in Siberië.
1 milj. km2 werd daar met lava bedekt. In vrij korte tijd (?).
Dankzij vrijkomende SO2 ontstond er zure regen, samen
met groot broeikaseffect.
In Zweden ontstond toen de Siljam-krater van 51 km. breed.
Veel landplanten verdwenen; ook bomen.
Grote hoeveelheden sediment werden afgevoerd naar zee met als gevolg algenbloei
en anoxie. Daarop volgde een periode van afkoeling dankzij
verlaging van het CO2 gehalte. Deze gebeurtenissen vonden alle plaats binnen
enkele tientallen jaren (?).
3. Extinctie tijdens Perm-Trias. 250 milj. jr. geleden.
Tussen Hercynische en Jurassische orogenese.
96% van de organismen sterft uit in de loop van 60.000 (?) jaar.
Grote bossen worden weggevaagd, samen met vele insectensoorten
De Siberische trappen ontstaan met 3 milj. km2 lava.
Dankzij de CO2 stijgt de temperatuur met 14 à 20o C.
De oppervlaktetemperatuur van het zeewater is ong. 40o C.
Samen met snelle erosie en algengroei ontstaat anoxie.
4. Extinctie tijdens Trias-Jura. 200 milj. jr. geleden.
Tussen Hercynische en Jurassische orogenese.
6 à 7 graden temperatuurstijging dankzij de
Centraal-Atlantische-provincie (CAMP).
Dit waren lavastromen in het Oosten van Zuid-
en Noord-Amerika en West-Afrika.
De hoeveelheid lava zou heel de VS met 400 m.
kunnen overdekken.
Dit gebeurde tijdens het ontstaan (??) van Pangea.
Of juist tijdens het eerste uiteengaan van Pangea?!
5. Extinctie tijdens Krijt-Tertiair. 66 milj. jr. geleden.
Door het treffen met een grote asteroïde van de aardbol in Mexico
sterft 75% van alle leven uit. Ook de Dinosauriers (zie hoofdst. 3.2.3).
Door stofstormen ontstaat er sterke afkoeling.
Vgl. de Alpiene orogenese.
In India vinden we de Deccan Traps;
bestaande uit 2 km. dikke basaltlagen dankzij vulkaanuitbarstingen.
In de omgeving een krater met een diameter van 40 km.
https://upload.wikimedia.org/wikipedia/commons/thumb/0/0a/India_Geology_Zones.jpg/260px-India_Geology_Zones.jpg
Het paarse gebied: de Deccan Traps. Denk hierbij aan het ontstaan van de Himalaya.
Eveneens een opvallende, vrij recente, gebeurtenis betreft het uitsterven van het grootste deel van alle haaiensoorten. Gerdurende een "oogwenk" van 100.000 (?) jaar. Haaien leefden al sinds 400 miljoen geleden jaar op de aardbol. Waardoor verdween aan het begin van het Mioceen 90% van de populatie?
Hoofdstuk 4 Belangrijkste vindplaatsen van fossielen.
De vindplaatsen van fossielen en de bijbehorende leefomstandigheden heb ik vooral ontleend aan het boek van Tirion Natuur; geschreven door Douglas Palmer.
4.1 Algemeen overzicht.
Vanaf het Precambrium t/m het Devoon bevinden alle vindplaatsen zich in ondiepe kustgebieden. Er waren wellicht haast eindeloos lange kusten rond de vele "continenten". Ik denk daarbij aan de situatie die bestond vanaf Pangea; de wereld vanaf de schepping!
Daar leefden vooral dieren (samen met algen en bacteriën) zonder harde delen van bijna alle nu bekende stammen van het dierenrijk: de Ediacara. Een geringe toenamen van stromend rivierwater of verhoging van het land was voldoende om door de toegenomen sedimentatie dit leven te bewaren als fossielen.
Dieren die zich daaraan konden onttrekken waren schelpdieren, brachiopoden, bodemvissen (agnaten), krabben, kreeften, zeesterren, wormen en Pikaia (de chordaat). Dankzij hun grotere bewegelijkheid.
Een nog veel sterkere stroom van sedimenten was nodig om echte vissen (straalvinnigen), nautiloïden, inktvissen, roggen, haaien, longvissen en sarcopterygiers (kwastvinnigen) voor eeuwen vast te leggen.
Die sedimenten ontstonden wellicht door de vroege processen van orogenese. Het moest allemaal in snel tempo gebeuren, vooral ook omdat deze fossielen vaak in grote aantallen bij elkaar worden gevonden. De verdwijning van deze dieren verliep heel snel in het warme kustwater. En er waren meer dan voldoende roofdieren en aaseters.
Tegelijkertijd groeiden op vele plaatsen op het land nabij de lage kusten levermossen, mossen, korstmorsen, sporenplanten (Cooksonia), varens, paardenstaarten en hogerop (dan praten we over de periode Carboon en Perm) bomen als Sigillaria, Lepidodendron, Calamites, Neuropteris (zaadvaren), Cordaïtes en naaldbomen.
De Hercynische orogenese (tezamen met continentverschuiving) was zeer waarschijnlijk noodzakelijk om deze complete flora te overweldigen en richting de kusten te drijven. Vervolgens vrij snel met dikke lagen sediment te overdekken om aldus bruinkool en diverse soorten steenkool te doen ontstaan.
De vindplaatsen liggen vaak richting kust of dichtbij berggebieden.
Op het land was intussen een rijk dierenleven aanwezig: Ichtyostega en Acanthostega (waterdieren met zwemstaart en kieuwen (en/of longen?) die ook het land opgingen), tetrapoden (vierpotige landdieren), hagedissen, kleine sauriërs, Dimetrodon,
 een Ichtyostega
een Ichtyostega
https://upload.wikimedia.org/wikipedia/commons/thumb/7/7f/Ichthyostega_BW.jpg/266px-Ichthyostega_BW.jpg
 Acanthostega
Acanthostega
https://upload.wikimedia.org/wikipedia/commons/thumb/a/a6/Acanthostega_BW.jpg/266px-Acanthostega_BW.jpg
 Dimetrodon
Dimetrodon
https://upload.wikimedia.org/wikipedia/commons/thumb/2/21/Dimetrodon_pair.jpg/250px-Dimetrodon_pair.jpg
In Permafzettingen is er sprake van grote riviervlakten zoals in Midden-Duitsland (toenmaals aan de kust gelegen) en Texas.
Vanaf het Trias nemen de sauriërs in aantal en grootte toe. Belangrijke vindplaatsen zijn dan:
- het Moskoubekken (rivieren vanaf de Oeral)
- kust van Antarctica (met grote coniferen-bossen)
- Zuid-Duitsland (nabij de Alpen met coniferen-bossen)
- Noord-Italië (ten zuiden van de Alpen, met vele zeereptielen)
- ten noorden van de Himalaya (rivieren, moerassen, meren met
bossen van palmvarens, ginkgo's, benettitales en coniferen)
- Noord-Amerika (nabij de Rocky Mountains, grote bossen met vele sauriërs)
- de kust van Zuid-Engeland (vele zeedieren en sauriërs)
- grote zeebekkens met soms diepe riffen.
Met overstromingsgebieden nabij bergen of kusten.
Vele terreinen in West-Europa, Zuid-Amerika, Noord-Amerika, China,
Australië, Nieuw-Zeeland en Antarctica
Vele fossielen zijn opgedolven in enkele savanne-gebieden; Mongolie en Noord-Amerika nabij de Rocky Mountains.
Tot aan het einde van het Krijt leefden vele reusachtige Dino's (lopend, vliegend, zwemmend). Daarnaast vogels, zoogdieren, geleedpotigen, bedektzadigen (angiospermen) enz. Meest opvallend was misschien wel Archaeopteryx tezamen met andere gevleugelde dino's die in ieder geval veren bezaten.
Na de tweede grote extinctie aan het einde van het Krijt komen we in de wereld die veel op de onze gaat lijken als we letten op de continentenverdeling rond de aardbol en de samenstelling van de enorme rijkdom aan planten en dierenleven. De wel of niet plaatsgevonden hebbende ijstijden (hoeveel?) bepaalden grotendeels de verhuizingen van planten en dieren.
Beroemd werden al de grotten waarin botfragmenten, werktuigen en rotstekeningen van hominiden en mensen werden ontdekt.
En wel in Spanje, China, Zuid-Afrika, Zuid-Amerika en het Rifgebied (de Alduvaikloof) in Afrika.
Op het eiland Flores werden grotten ontdekt met hominiden die dwerggroei vertoonden en een kleine schedelinhoud (650 cc). Ze maakten jacht op dieren met zeer eenvoudige wapens. Waren het "apen" of "mensen"?
4.2 Vindplaatsen met grote hoeveelheden fossielen.
West-Europa.
Eurazië en Afrika.
Noord- en Zuid-Amerika.
Verklaring: de nrs. geven de vindplaatsen zoals vermeld in
Tirion Natuur. De vindplaatsen genoemd door Velikovsky zijn aangegeven
met de letters A t/m H.
De bergen ontstaan tijdens de Caledonische orogenese
zijn weergegeven met kleur oranje.
De bergen ontstaan tijdens de Hercynische orogenese
zijn weergegeven met de kleur groen.
De bergen ontstaan tijdens de Alpiene orogenese
zijn aangegeven met de kleur
rood. Overige berggebieden zijn aangegeven met de kleur bruin.
Deze vindplaatsen hieronder in volgorde van ontstaan: (uitgezonderd de laatstgenoemde: Monte Bolca, 50 milj. jr. geleden):
3. Noord-Korea (Chengjiang): 150 soorten in ondiepe zeekust. Bijna alle grote groepen dieren zijn aanwezig van sponzen en wormen t/m vertebraten (gewervelden); waaronder arthropoden (trilobieten e.a.).
4. Burgess Shale (Canada): onderaan een onderzees klif. Een overvloed aan fossiele soorten, vooral arthropoden.
5. Trenton (westkust van de VS): 500 families van zeedieren. koraalriffen, slangsterren, inktvissen, nautiloïden enz.
12. Hunsrück (Duitsland): sponzen, koralen, brachiopoden, crustaceeen, echinodermen, arthropoden, inktvissen, zeespinnen en haaien. Van de vissen: agnatha, placodermen, en longvissen (sarcopterychiers).
13. Gogo (west Australisch kustgebied): actinopterygii (beenvissen), sarcopterygii (kwastvinnigen), placodermen en haaien.
14. Miguasha (kust tgo. New Foundland): 1000-den fossielen; 20 soorten vissen waaronder sarcopterygii, actinopterygii en longvissen. Veel restanten van landplanten vermengd met arthropoden.
En Archaeopteris. Ook varenachtige bladeren en een houten stam van het gymnosperme type. De eerste boomvorm op aarde? 18 m. hoog.
19. Bear Gulch (VS, ten oosten van de Rocky Mountains): 100 soorten vissen (4800 stuks, snel begraven) en 5 soorten kraakbeenvissen, waaronder haaien.
20. Mazon Creek (VS, ten zuiden van de grote meren): 150 soorten arthropoden, waaronder libellen, schorpioenen en duizendpoten. En amfibieën. Van de landplanten: wolfsklauwen, zaadvarens en paardenstaarten.
22. Nyrany (Tsjechie): 150 soorten arthropoden, grote insecten (libellen), 30 soorten vissen en 20 soorten tetrapoden, sauriërs en amnioten.
26. Glossopteriden-wouden (oost Australië): 300 m. dikke steenkoollagen, Dus wolfsklauwen, zaadvarens, coniferen, paardenstaarten.
27. Karoo-bekken (Zuid-Afrika): 70 soorten tetrapoden waaronder cynodonten, kleine tot grote planten- en vleeseters.
32. Monte San Giorgio (noord Italië): 100-den grote zeereptielen. 1000-den vissen en schelpdieren.
33. Ferganavallen (Kirgizie): coniferen, palmvarens, ginkgo's, benettitales. Vele insecten, waaronder titanoptera met een vleugelspanwijdte van 40 cm.
34. Valle de Luna (Zuid-Amerika): 40 m. reusachtige coniferen en grote sauriers.
36. Hayden Quarry (VS ten oosten van de Rocky Mountains): 1300 vertebraten, en (bij Ghost Range) 100-den skeletten van Coelophysis (de snelste jagers onder de dino's).
37. Lyme Regis (zuiden van Engeland): grote hoeveelheden fossielen nu aanwezig in musea over de gehele wereld. Vele grote zee- en lucht sauriërs (Ichtyosauriers, Plesiosauriers, Pterosauriers) samen met vissen en haaien.
38. Holzmaden (zuiden van Duitsdland): 500 complete skeletten van Ichtyosauriers en 70 teleosaurische krokodillen.
41. Tendaguru (oostkust van Afrika): 200 ton fossielen weggesleept, waaronder zeer grote dino's zoals Brachiosaurus van 22 mm. En: Ramphorynchus, de grote "vogel".
42. Guimarota (kust van Portugal): 10.000-den terrestrische en aquatische fossielen. Schelpdieren, arthropoden, dino's (b.v. Ramphorynchus; geen echte dino), schildpadden, krokodillen, vogels en 26 soorten zoogdieren en tanden van Archaeopteryx.
43. Solnhoven (Duitsland ten noorden van de Alpen): Archaeopteryx, Rhamphorynchus en Pterodactylus. 1000-den fossielen en 600 soorten zee- en landdieren.
44. Morrisonformation (VS oostelijke Rocky Mountains): 1.5 miljoen km2. 44 grote dino's met o.a. Diplodocus, Camarosaurus, Apatosaurus, Camptosaurus, Stegosaurus, Allosaurus
Dry Mesa Dinossaur Quarry (VS Colorado): 23 soorten dino's waaronder Supersaurus en Megasaurus. Samen met Pterosauriers, krokodillen, schildpadden, amfibieën, longvissen en vele zoogdieren.
45. Bernissart (kust Belgie): 30 Iguanodonten, en Ornitocherus, een Pterosaurier. .
46. Liaoning (China): "vliegende" dino's en dino's met veren die dienden tot verwarming of lokken van partner(?) Verder zoogdieren en vogels. 1000-den fossielen waaronder bloeiende planten (angiospermen).
48. Crato Formatie (oostkust van Brazilië): 100-den insectensoorten, 11 soorten spinnen, 9 vissoorten, Pterosauriers, veren van sauriërs en vogels samen met Nymphaeales.
49. Dinosauer Cove (Australië): 1000-den botten van sauriërs.
51. Neuquén (Zuid-Amerika): reuzesauriërs (o.a. Gigantosaurus) en 1000-den nesten van sauriërs. Vele tetrapoden, krokodillen, slangen en zoogdieren.
52. Ukhan Tolgod (China): 200 dinoskeletten waaronder Protoceratops en Oviraptor. Ook 1000-den zoogdierskeletten (vele compleet) en 1400 hagedissen.
53. Judith River (Canada, zuid Alberta ten oosten van de Rocky Mountains): 100-den dino's. Veel angiospermen waaronder eikenbomen, 20 soorten zoogdieren en de vogel Quetzalcoatlus
55. Hell Creek (VS ten oosten van de Rocky Mountains): 100 soorten zoogdieren (3x zoveel als dino's) samen met Tyrannosaurus rex, Triceratops en 100-den soorten angiospermen.
56. Crazy Mountains (VS ten oosten van Rocky Mountains): marsupialen (buideldieren), multituberculaten ("kaagdieren"), placentalen (88 geslachten).
58. Green River (VS ten oosten van de Rocky Mountains): 65.000 km2 en een laag van 2 km. meersedimenten. Een miljoen fossielen verzameld: schimmels, planten, insecten, weekdieren, vissen, hagedissen, slangen, schildpadden, krokodillen, vogels en zoogdieren (paarden, tapirs, neushoorns en Uinthaterium (zo groot als een olifant). En een vleermuis!
59. Abbey Wood (zuiden van Engeland): zoogdieren, vleermuizen, primaten, paarden, pantodonten, pantolestiden, apatotheria, creodonten, condylarta en artiodactyla (hoefdieren). Samen met 300 soorten angiospermen.
60. Messel (Duitsland): vogels, vissen, krokodillen, schildpadden, salamanders, kikkers, kevers (en vele andere soorten insecten). Ook vele hagedissen, leguanen en slangen. Tot de vogels behoorde o.a. de niet-vliegende Diatryma, nu Gastornis.
https://upload.wikimedia.org/wikipedia/commons/thumb/3/39/PSM_V21_D483_Skeleton_of_a_gastornis_eduardsii.jpg/260px-PSM_V21_D483_Skeleton_of_a_gastornis_eduardsii.jpg
Deze dieren samen met vele soorten planten waaronder dennen, beuken, kastanjes en eiken.
Zoogdieren waren: opossums, insecteneters, viseter, schubdieren, miereneter, carnivoren, knaagdieren, vleermuizen, evenhoevigen en 4 soorten "paarden".
62. Fayum (Egypte, Nijldelta): jacana's, flamingo's, reigers, ooievaars, aalscholvers, visarenden, krokodillen, zeekoeien en schildpadden. Zoogdieren: neushoornachtigen, hyaenodonten, primaten en olifantachtigen.
63. Florissant (VS Colorado): 1500 soorten insecten; zeer gave vlinders. Vele bloemplanten.
65. Riverleigh (noorden van Australië): marsupialia, Neohelos ("nijlpardachtige"), buidelleeuw, kangoeroes, krokodillen, slangen, opossums, (loop)vogels, wallaby's en buidelmollen.
Daarnaast vele waterdieren.
67. Silawikformaties (noorden van India): 2-5 km. dikke lagen. Fossielen als dikke aggregaten; 40.000 exemplaren. 13 orden zoogdieren waaronder artiodactyla. Vogels, apen, neushoorns, herten, hyaena's enz. En alle soorten waterdieren.
68. Djurabwoestijn (midden van Afrika): Sahelanthropus, vele soorten waterdieren, amfibieën, knaagdieren, olifanten, paarden, runderen en apen.
70. Sterkfontein (grotten in Zuid-Afrika): Hominiden (Australopithecus africanus, 500 ex.) samen met runderen, antilopen, hyaena's, sabeltandkatten, leeuwen, luipaarden en arenden. De roofdieren en aaseters leefden in grotten?
78. Magadan (oostelijk kustgebied van Siberië): Dieren gevangen in permafrost. Mammoeten, wolven, neushoorns, bizons en paarden.
79. La Brea (VS noordelijk Californië): 166.000 botten van sabeltandtijger Smilodon. Bewaard in teerlagen waarin ook planteneters, wolven, coyotes en roofvogels werden gevangen. In totaal 600 soorten! 1 miljoen beenderen van 58 zoogdiergeslachten: kleine knaagdieren, insectivoren, vleermuizen, hazen, bizons en leeuwen.
Overige soorten: 138 vogels, 24 reptielen, 6 amfibieën, 3 vissen, 56 mollusken, 168 arthropoden, 80 plantentaxa (groepen).
86. Monte Bolca (Italië ten zuiden van de Alpen): 500 soorten land- en zeeorganismen. 250 soorten vissen,
4.3 Opvallende vindplaatsen en gebeurtenissen.
1. Vele vindplaatsen bevinden of bevonden zich aan de oevers van de zeeën of zeer grote meren.
Dit geldt voor de nrs.
54, 45,59, 37,7,7,10,9,18,11,35,8,72,42,12,25,60,22,31,38,43,40,80,32,86,29,27,28,78,47,46,73,52,61,39,3,67,50,65,13,2,76,49,26,30,62,66,71,69,41,70,74,7,23,15,1,21,5,16,20,79,48,34,82. In totaal 57. Dat is 70%.
Een aantal van de vindplaatsen bevindt of bevond zich aan de rand van de orogenese-zones. Gedeeltelijk valt dat samen met kustgebieden. Dit zijn de nrs. 9,10,11,18,31,38,43,40,32,86,33,28,67,50,65,76,49,84,40,71,69,70,4,53,55,19,56,57,58,85,44,63,36,83,6,51,34. In totaal 37. Het totaal van beide reeksen is dus meer dan 100%.
Het is dan ook duidelijk dat er slechts enkele gebieden zijn die geheel buiten de kusten of gebergten vallen. Het zijn de nrs. 27,64 en 68.
2. Als we aannemen dat de processen van continentverschuiving, orogenese en vulkanisme (samengaand met vaak aardbevingen en tsunami's) zich hebben voltrokken binnen
betrekkelijk korte tijd dan is het niet moeilijk om al de aardperioden van precambrium tot en met het holoceen samen te brengen in 8000 à 10.000 jaar. Als argument kan in ieder geval dienen dat het invriezen van enorm grote dieren als mammoeten en neushoorns in zeer korte tijd moet hebben plaats gevonden. Omdat vele weefsels die snel vergaan door bacteriën en schimmels of gegeten worden door roofdieren en aaseters anders niet waren bewaard gebleven.
3. Het voorkomen van de midatlantische ruggen op de bodem van alle grote wereldzeeën maakt dat de processen die de verplaatsing van de continenten veroorzaakten vrij snel hebben kunnen verlopen. Tijdens die processen verwisselden Noord- en Zuidpool een aantal keren van plaats. Oftewel het aardmagnetische veld keerde een aantal malen om. Dat zal hoogstwaarschijnlijk snel gebeurd zijn.
Geleerden hebben vastgesteld dat eens in de 200.000 tot 3000.000 jaar het magnetische veld omkeert. Dus ruwweg 3 tot 8 keer per miljoen jaar! Alan Cooper van het museum in Adelaide meent te hebben berekend dat dit de laatste keer gebeurde 42.000 jaar geleden. Dit dankzij de koolstofdatering in jaarringen van de fossiele kauriboom (Agathis australis). Tijdens zo'n omkering van het veld verzwakt de magtnetosfeer. Tot wel 6% van de normale waarde. Dit heeft ingrijpende gevolgen:
- verhoging van de kosmische straling
- toename radioactief C-14
- toename ioniserende straling
- beschadiging van de ozonlaag
- toename ultraviolette straling
- hoge temperaturen en andere extreme weersomstandigheden
- uitstervingsgolven van o.a. gigantische zoogdieren en Neanderthalers.
Het meest bijzondere is dat er gedurende lange periode van het Krijt (ong. 40 miljoen jaar) geen omwisseling van het magneetveld plaats vond. (zie Earth History and Paleogeography, pg. 4: CNS = Cretaceous Normal Superchron). Ik veronderstel: Dit was de periode vanaf de eerste scheppingsdag.
Dat de platen nu nog slechts enkele mm's of cm's per jaar verschuiven is heel vanzelfsprekend.
Breuklijnen en verschuiving van de platen.
https://f.jwwb.nl/public/n/q/k/temp-qdavnyuyfzafoznhpmxa/breuklijnen.jpg
4. Het ontstaan van fossiele organismen is heel onwaarschijnlijk. Onder "normale" omstandigheden ontstaan er geen of nauwelijks fossielen. Dieren groter dan 50 cm. of 1 m. zullen altijd opgegeten worden. Ook de botten; door aaseters. En alle dieren vergaan vrij snel door schimmels, bacteriën en kleine organismen in de grond. De verzamelingen vrij grote organismen op vele plekken op aarde kunnen daarom onmogelijk hun oorzaak vinden in langdurige sedimentatie- en versteningsprocessen.
Om dit laatste toch aannemelijk te maken wordt er gespeeld met fantasieën als: zuurstofloze wateren, snelle afkoeling van water, verzuring van moerassen, meegesleurd worden door rivierstromingen enz.
5. Als gebergtevorming vrij snel verloopt kunnen geweldige stortvloeden van water met aardlagen (zand, klei, modder) enorme oppervlakten bedekken en veel wat leeft meesleuren. Ook boomstammen en struiken. Dieren en planten worden dan niet netjes gesorteerd maar kris kras door elkaar gesmeten en verbroken in vele stukken. Ook roofdieren samen met planteneters.
Als orogenese samen ging met veel vulkaanuitbarstingen, aardlagenoverschuivingen en zeer snelle temperatuurdaling zijn vele merkwaardige verschijnselen rond vindplaatsen van "hopen" fossielen begrijpelijk.
Velikovsky schreef zijn boek Earth in upheaval in 1956.
Velikovsky verdedigde in dit boek opvattingen die lijnrecht ingingen tegen de toenmalige en huidige theorieën over ontstaan van aardlagen, ijstijden, gebergtevorming en fossilisatie.
Vooral het tempo waarmee de geologische processen plaats zouden hebben gevonden betwist hij in hoge mate. En daarmee de krachten waardoor deze enorme gebeurtenissen plaats vonden.
Ongetwijfeld waren bepaalde opvattingen en uitgangspunten van Velikovsky niet vrij van rijke fantasie en overdrijving. De feiten die hij met altijd uitvoerige bronvermelding ter tafel brengt zijn steeds de moeite van leren kennen waard. Daarom hier een overzicht van door hem vermelde vindplaatsen van fossielen.
(Op de kaarten zijn deze aangegeven met de letters A t/m/ H)
A. Het Fairbanksdistrict in Alaska: een enorm aantal beenderen in een laag sediment van 45 m. Beenderen van miljoenen dieren: mammoeten, bizons en paarden. Met grote hoeveelheden bomen die in stukken gebroken zijn evenals de botten van de dieren.
B. Liakhof eilanden, 9600 km. binnen de poolcirkel. Het eiland lijkt wel opgebouwd uit beenderen van mammoeten en slachttanden van olifanten.
C. Eilanden van Stolbovoi en Belkor: zelfde situatie als hierboven genoemd. Daarbij ook neushoorns.
D. Old Red Sandstone in Schotland. Half Schotland bestaat uit dit gesteente; 2650 m. hoog. In deze zandsteen miljoenen vissen en andere zeedieren. Vaak complete skeletten, dicht opeengepakt.
Vergelijkbare situaties in Bolca (Italie), de Harz, Thüringen, Saarbrücken, Solnhofen (Duitsland), diverse plaatsen in Zwitserland en in Aix en Provence.
E. Victoria Cave in Yorkshire: ontelbaar veel mammoeten, neushoorns, nijlpaarden, bizons, hyena's e.a. Vergelijkbare vondsten in Wales.
F. Norfolk: grote en dikke afzettingen van bomen met 60 diersoorten en 68 plantensoorten. Organismen afkomstig van koude, gematigde en tropische gebieden.
G. Noord-China bij Peking: onnoemelijk veel beenderen in spleten en grotten. Dieren van de toendra, steppen en oerwouden.
H. Vero en Melbourne in Florida: aardewerk door mensen gemaakt. Samen met dierenleven uit het Pleistoceen. O.a. de sabeltandtijger Smilodon.
Rotskloven op vele plaatsen in Engeland, Wales, Centraal Frankrijk, Lyon, Dyon, de franse Middellandse zeekust, Gibraltar, Corsica, Sardinië en Sicilië. Daarin worden vele soorten fossielen gevonden van groot tot klein; zowel van planten als dieren. Veelal versplinterd en verbroken.
Als we alle vindplaatsen die beschreven zijn in het boek 'Trition Natuur' in volgorde bekijken valt op dat er pas 3.5 miljard jr. geleden de eerste tekenen van leven op aarde ontstaan. Vier en een half miljard jr. daarvoor helemaal niets! Gezien vanuit de bijbelse geschiedenisopenbaring is dit tyotaal ongeloofwaardig!
Na die 4 miljard jr. is er ong. 3 miljard jr. lang nog bijna niets. Slechts de Stromatolieten komen tot leven aan de kusten van West-Australië: Algen en cyanobacteriën.
Twee miljard jr. geleden werden er minimale structuren gevonden in Rusland, Canada en West-Australië waarvan het nog maar de vraag is of het organismen waren.
Vanaf 575 miljoen jr. worden er fossielen bewaard van kustgebieden met ondiepe zeeën. Vaak in de buurt van bergketens. Of moerasgebieden, rivieren en meren. Dit gaat door, ook na de grote extinctie einde Perm, tot 20 miljoen jr. geleden.
Deze vindplaatsen bevinden zich rondom de aardbol: Omgeving New Foundland, Zuid-, Noord- en Oost-Australië, Noord-Siberië, kusten van China, West- en Oost-Canada, Oost- en West- Amerika, gebied van de grote meren in de VS, Zuid-Amerika, Zuid-Afrika, Whales, Duitsland, Groenland, Wolga-bekken, Frankrijk, Antarctica, Engeland, Schotland, Portugal, Brazilië, Nederland.
Vele van de vindplaatsen lagen in de nabijheid van berggebieden. Vooral die ten oosten van de Rocky Mountains, ten noorden en ten zuiden van de Alpen, ten noorden en ten zuiden van de Himalaya, ten oosten van de Andes. Gebergten die ontstonden door de Alpiene orogenese of berggebieden ontstaan door de Hercynische- of Caledonische orogenese.
Vindplaatsen die verband houden met (korte?) perioden van orogenese gaan vaak gepaard met de vorming van steenkool.
V.b. Joggins (VS) 900 m. dik, 45 steenkoollagen
Nyrany (Z.O. Duitsland) In de bossen leven sauriërs en vele andere dieren
Glossopteridenwouden in Australie
Fergano (Zuid-Rusland) nabij de bergen in Kirgizië
Guimarato (Portugal) met sauriërs, zoogdieren enz. enz.
Morrison Fomation (Rocky Mountains) met reuze-sauriërs en vele andere diersoorten.
Bijzonder vind ik dat in de steenkoolformaties veelal allerlei soorten insecten worden gevonden. Dit zijn dieren die normaliter onmiddellijk dienen als voedsel voor allerhande predatoren!
Hoogst merkwaardig: de honderden (of duizenden) ingevroren mammoeten (en sommige andere grote dieren) in de permafrost van Siberië.
Welhaast nog vreemder zijn de grote hoefdieren en roofdieren, samen met roofvogels, knaagdieren, vleermuizen en hazen, insecten en vogels die omkwamen in de teervelden van La Brea. Het fabeltje dat hieromheen verteld wordt is ongelofelijk merkwaardig. Dieren lopen niet per ongeluk in sloten of over te dun ijs. Ze vallen zelfs meestal niet uit bomen. Vogels, vleermuizen en insecten kunnen niet omkomen in teervelden. Een ramp van heel bijzondere aard moet in die omgeving hebben plaats gegrepen waardoor deze totaal verschillende dieren in de daar aanwezige teervelden werden neergesmeten.
Eenzelfde onwaarschijnlijk verhaal wordt vaak verteld over de vele vindplaatsen van vliegende en zwemmende grote sauriërs en moderne vissen, insecten, vogels en zoogdieren.
Opm. de gehanteerde perioden dienen slechts als hulpmiddel om een veronderstelde volgorde aan te geven.
Hoofdstuk 5 De Dinosauriërs.
Steve Brusatte vertelt hierover in zijn boeiende boek De opkomst en de ondergang van de Dinosaurus (2018).
Brusatte is een groot kenner van de Dino-wereld. Hij heeft meegedaan aan vele veldonderzoeken en heeft vele skeletten en beenderen in musea over de hele wereld onderzocht.
Zijn boek is een goed hulpmiddel om van dino's te gaan houden. Behulpzaam daarbij was het boek van Tirion natuur Evolutie met geweldig mooie illustraties. Met daarnaast De oerwereld van de Dinosauriërs (eveneens van Tirion Natuur, 1993). En niet te vergeten: Encyclopedia of Dinosaurs o.l.v. Peter Dodson (1990).
In 2015 beschreef Brusatte samen met een collega het skelet van Zhenyuanglong suni. In China.
Het skelet vertoonde opvallende kenmerken:
- het had lichte, holle beenderen
- magere poten als van een reiger
- een slank skelet
- bedekt met "veren".
Veren tussen aanhalingstekens omdat ze borstelig waren en op hoofd- en nekharen leken. Lange "veren" op de staart en elkaar bedekkende "veren" op de voorpoten. Dus ze hadden "vleugels".
Let op de bijzondere positie van dit skelet! Dit komt heel vaak voor.
https://upload.wikimedia.org/wikipedia/commons/thumb/7/7b/Zhenyuanlong.jpg/266px-Zhenyuanlong.jpg
Op de conclusie van de schrijver: "Deze dinosauriër leek precies op een vogel" valt m.i. wel wat af te dingen.
Daar kom ik aan het slot van dit hoofdstuk wel op terug.
Hiermee is de schrijver aangekomen bij de afsluitende periode van zijn onderzoek. Dit om meteen zijn
grote interesse voor de afstammingsidee van de vogels uit dino's vast te stellen.
Ong. 340 milj. jr. geleden, tijdens het onder-Carboon, werden de eerste amniotische eieren op het land gelegd. Probeer je in te denken welk een kolossaal veranderingsproces dit betekent!
Daarvoor nog maar een keer de bouw van een vogel- of reptielen ei:
1. kalkschaal
2. schaalvlies
3. binnenste schaalvlies
4.
hagelsnoer5. buitenste eiwit
6. middelste eiwit
7. dooiervlies
8.
dooier9. kiemvlek (of vormingsdooier)
10. donkere eigeel
11. klare eigeel
12. binnenste eiwit
13. hagelsnoer
14. luchtkamer
15. cuticula
Hoeveel mutaties zouden hiervoor nodig zijn??
Denk daarbij ook aan:
- de bouw van de voortplantingsorganen
- het (broed)gedrag van de dieren
- de voortplantingsrituelen
- de zintuigen
- de ontwikkelingen van het jong in en buiten het ei
- de hersenen.
Het alleroudste reptiel is niet bekend. Dus er bestaat slechts een voorouder in de fantasie!
In de tijd van de Dinosauriërs leven ook de Zoogdieren.
Deze twee groepen beconcurreren elkaar niet omdat zoogdieren een totaal andere bouw en wijze van voortplanting kennen. Denk aan baarmoeder, placenta, melkklieren, beharing enz. enz.
Binnen bepaalde ecosystemen hebben de zoogdieren altijd een enorme voorsprong als het gaat om zich handhaven t.o.v. reptielen. De reusachtige grootte van vele sauriërs speelt hierbij nauwelijks of geen rol.
Toepassing van macht en geweld speelt geen belangrijke rol in het dierenrijk.
5.1. Gedurende het Perm.
Tijdens het Perm verandert het klimaat. Pangea had eerst in het geheel een vochtig en warm klimaat zonder seizoenen. Geen ijsgebieden bij de polen. Zeestromen zorgden voor een gelijkmatige verdeling van de temperatuur. Later verandert dit. Er ontstaan natte en droge perioden en koude en warme (hete) seizoenen. Rond de evenaar wordt het warmer en richting de polen kouder.
Vele amfibieën en reptielen leven in de dichtbegroeide wereld van de kusten en overstromingsgebieden. Kleine insectenetende reptielen trekken hogerop de drogere gebieden in.
In de laaglanden "experimenteren" de reptielen met nieuwe eigenschappen en ontstaan vele nieuwe groepen.
Als eerste de
Anapsida: de schedel achter de oogkassen is helemaal gesloten.
Dat heet 'primitief'. Geen enkel organisme is ooit primitief gebouwd. Bij nader onderzoek blijkt altijd de meest perfecte bouw om te kunnen overleven. Zowel bij (cyano-)bacteriën, schimmels, wormen enz. enz.
De Anapsida zijn einde Perm uitgestorven. Schildpadden kunnen als Anapside beschouwd worden. Het kan ook zijn dat hun schedelopening is dichtgegroeid. Ze ontstonden in het Trias.
Als tweede de Synapsida: met één extra opening achter de oogkassen. Hiertoe behoorden de zgn. zoogdierreptielen (o.a. de Cynodonten) en de huidige zoogdieren.
Als derde de
Diapsida: achter de oogkassen bevinden zich twee openingen.
Hiertoe behoren de krokodillen, hagedissen en slangen samen met alle uitgestorven dino's. Ok de vogels worden hiertoe gerekend, hoewel ze beide openingen missen. Op grond van de evolutiegeschiedenis moeten beide openingen zijn dichtgegroeid. Dit type omgekeerde redenering komt nogal eens voor! Zo wordt de theorie schijnbaar gered!
Als vierde de Euryapsida: er is nog slechts één opening aanwezig doordat de bovenste opening is verdwenen(?) Dit was het geval bij de Ichtyosauriers en de Plesiosauriers.
De eerste
synapsida zouden de
Pelycosauriers geweest zijn. Ze waren omnivoor en hun nazaten werden herbivoor of carnivoor. Ze werden groter en ontwikkelden wel of geen rugzeil. Die rugzeilen waren wellicht nuttig voor de temperatuurregeling of voor pronk- en dreiggedrag.
https://upload.wikimedia.org/wikipedia/commons/thumb/f/fe/Ophiacomorphs2.jpg/266px-Ophiacomorphs2.jpg
Pelycosauriers.
Nu leert de theorie dat de Dinosauriërs niet uit de eerste reptielen ontstaan zijn. Nee, uit de reptielen ontstonden de zoogdierachtige reptielen, de Therapsiden. En daaruit vervolgens de Dinosauriërs.
Uit zeer vroege reptielen (synapsida) ontstaan de voorlopers van de zoogdieren, de Cynodonten.
De Pelycosauriers speelden hierin een belangrijke rol.
In een groeve bij het Orlando-meer in Oklahoma worden vele skeletten van deze dieren gevonden: Dimetrodon, samen met haaien, longvissen en grote amfibieën. De laatste zijn tot grote hopen op elkaar geperst.
https://upload.wikimedia.org/wikipedia/commons/thumb/2/21
/Dimetrodon_pair.jpg/250px-Dimetrodon_pair.jpg
Beroemde v.b. van amfibieën uit die tijd waren Seymouria en Diadectes. Ze worden beschouwd als overgangsvormen tussen reptiel en amfibie. Diadectes had snijtanden voorin en maaltanden achterin de bek. Ze worden wel gerekend tot de Reptiliomorpha. De grote vraag is natuurlijk of ze eieren op het land legden en dus amnioot waren. Diadectes had de aanleg voor een verbeend gehemelte. Belangrijk voor de ademhaling. Dit komt ook voor bij latere reptielen maar is op zichzelf geen argument voor ontwikkeling in de richting van de reptielen. Evenmin als de maalkiezen trouwens.
 Seymouria
Seymouria 
Diadectes.
https://upload.wikimedia.org/wikipedia/commons/thumb/1/1d/Bromacker2_copy.jpg/260px-Bromacker2_copy.jpg
 Watongia.
Watongia. Bron Wikipedia.
Watongia komt in beeld als het gaat om een overgang tussen de Pelycosauriers en de Therapsiden.
Watongia was een vleeseter. De Therapsiden worden einde-Perm de overheersende groep. Ze komen ook voor op de hogere breedten, tot 700 NB.
Deze 'proto-zoogdieren' hadden de volgende zoogdierachtige kenmerken:
- een meer opgerichte houding met poten rechter onder het lichaam
- ze waren misschien levendbarend en zoogden hun jongen
- sommige hadden misschien een vacht van haren
- ze hadden uitstekende oorschelpen aan hun hondachtige kop met natte
neus, snorharen, soepele lippen en uithangende tong bij uithijgen.
- misschien een vermogen tot endotherme temperatuurregeling.
Bij veel van het bovengenoemde lijkt me de wens de vader der gedachten. Verreweg de meeste zijn onwaarneembaar/onbewijsbaar. Alleen de eerstgenoemde heeft werkelijkheidswaarde en is van belang voor een eventuele evolutionaire ontwikkeling.
Intussen blijft het onbegrijpelijk dat zoogdieren met hun enorme voorsprong voor wat betreft hun functionele lichaamsbouwbouw en verbazingwekkende voortplantingsmogelijkheden bijna met uitsterving werden bedreigd na miljoenen jaren leven op aarde. Na het Krijt kwamen ze wonderwel tot een overheersende verspreiding over alle gebieden op aarde.
Onder de Therapsiden in het Perm waren er dieren die hun heil zochten in terugkeer naar het water. Deze dieren moeten eierlevendbarend zijn geweest omdat ze geen amnioten waren. Bij deze terugkeer was er sprake van analoge ontwikkeling bij diverse groepen. Vgl. de huidige zeehonden, walvissen, vinvissen etc.
https://upload.wikimedia.org/wikipedia/commons/thumb/c/c7/Mesosaurus_BW.jpg/266px-
Mesosaurus_BW.jpg
De grote Dinocephalia komen tot wereldwijde dominantie maar sterven in het einde van het Midden-Perm uit. Niemand weet waardoor.
De Dicynodonten zijn tegen einde-Perm over de hele aarde verspreid: Zuid-Afrika, Zuid-Amerika, India, China, Rusland en Schotland.
Ze hebben een schildpadachtige snavel en twee slagtanden. Ze variëren in grootte van mol tot olifant. Er zijn honderden soorten. Bovendien zijn ze misschien
endotherm.https://upload.wikimedia.org/wikipedia/commons/thumb/d/d2/Placerias.jpeg/266px-Placerias.jpeg
Aan het einde van het Perm vindt het volgende plaats:
alle Gorgonapsida sterven uit,
bijna alle Therocephalia sterven uit,
vele Dicynodonten sterven uit,
alle
Pareiasauriers sterven uit. De laatste waren planteneters en behoorden tot de Anapsida. De Pareiasauriers behoren tot de
Cotylosauriers die beschouwd worden als de stamgroep van alle reptielen. De Cotylosauriers op hun beurt zouden zijn voortgekomen uit de
Labyrentodonten de belangrijkste groep van grote amfibieën.
https://upload.wikimedia.org/wikipedia/commons/thumb/5/58/
Pareiasaurus_serridens.jpg/266px-Pareiasaurus_serridens.jpg
De heerschappij van de Therapsiden wankelt maar herleeft in het Trias, door de opkomst van de diapside reptielen. De Archosauriers verschijnen op aarde. Daaruit ontstaan de Dinosauriërs
In het gebied van het Heilige Kruisgebergte in Polen worden duizenden pootafdrukken van hagedissen, amfibieën en zoogdieren samen met die van sauriërs gevonden.
Gedurende het Perm (250 milj. jr. geleden) leefden hier:
- salamanders zo groot als honden,
- Pareiasauriers,
- Dicynodonten (waaruit de zoogdieren ontstaan) zo groot als varkens; met vacht. Waren ze
warmbloedig?
Tegen het einde van Het Perm ontstaan er km's lange scheuren in de aardkorst van Noord- en Centraal-Azië. Miljoenen km
2 worden met lavastromen bedekt:
de grote basaltvlakten in Siberië. De coniferenwouden sterven af door de enorme stof- en gaswolken en de zure regen. Natuurbranden en modderstromen vagen de bossen weg. Het CO2 -gehalte stijgt enorm en de temperaturen lopen op.
Het zeewater wordt zuur en O2 arm. Schattingen geven aan dat 90% van alle organismen de dood vindt.
Er zijn daarna geen bloemplanten (angiospermen) meer (?).
5.2 Gedurende het Trias.
Opm.: Radiometrische datering (van K naar Ar) is alleen mogelijk als vloeibaar materiaal afkoelt naar vast gesteente; dus verandert in basalt of graniet. Van zand- en kleisteen kan met die methode niet de ouderdom worden bepaald. Een oplossing voor dit probleem is mogelijk als een sedimentgesteente zich bevindt tussen 2 vulkanische gesteenten. Soms kan de bepaling ook plaats vinden dankzij afzetting van kristallen in het sediment. Maar dit is een erg dure methode.
Binnen het Trias zijn er erg weinig dateringen!!
De eerste echte Dino werd gevonden in Argentinië. In de Valle de la Luna, dicht bij de Andes.
Hier werden enorme hoeveelheden botten en skeletten opgegraven van Herrerosaurus.
Eveneens de omnivoor
Eoraptor.
Deze dieren leefden temidden van:
Zoogdieren, Therapoden (vleeseters)
, Sauropoden (de grote, zware 'langnekken')
en Ornithischia (planteneters)
.
Het blijft vergeefs zoeken naar voorouders bij al de genoemde groepen!
Tijdens het Trias komen we in een wereld van hoge bergketens die ontstaan door bewegende landdelen. Er zijn geen ijskappen; geen poolgebieden. Er bestaan geen echte winters en de tropen zijn erg warm. Tussen 300 NB en 300 ZB vormen zich enorme woestijngebieden. Er ontstaan vele nieuwe soorten planten en dieren (?) zoals vroege zoogdieren, amfibieën en reptielen. Of worden die ontdekt in nieuw ontstane ecologische niches?
In Portugal, de Algarve, bevindt zich een 50 cm dikke kleisteenlaag vol met botten, liggend in een chaotische hoop. Honderden reptielen, schildpadden, salamanders, hagedissen, krokodillen, vele soorten insecten en zoogdieren.
Maar er waren geen Dino's in de drogere gebieden van Spanje, Portugal, Marokko en de oostkust van Noord-Amerika. Wel in Brazilië, India en Argentinië.
Ecologische omstandigheden bepaalden blijkbaar grotendeels het vinden van fossiele diersoorten!
In vochtiger gebieden van het Noordelijk Halfrond, in Duitsland, Frankrijk en Zwitserland worden massagraven ontdekt met o.a. Plateosaurus. Er zijn wel 50 vindplaatsen met vele skeletten temidden van andere dieren als schilpadden, krokodillen en kleine zoogdieren.
https://upload.wikimedia.org/wikipedia/commons/thumb/b/b2/
Plateosaurus_trossingensis.jpg/260px-Plateosaurus_trossingensis.jpg
Het lijkt er op dat Dino's zich na verloop van tijd verplaatsten naar drogere en zelfs woestijnachtige gebieden. In Noord-Arizona en New Mexico met rode zandsteen en kleisteen vinden we de 500 m. dikke Chinle Formation. Gedurende het laat-Trias ontstond hier in het Forest National Park 1000-den gefossiliseerde bomen. Eveneens een massagraf van 100-den skeletten van Coelophysis. Ze zijn nu verspreid over vele musea over de hele wereld.
https://upload.wikimedia.org/wikipedia/commons/thumb/8/80/
Coelophysis_size.jpg/260px-Coelophysis_size.jpg
In dezelfde omgeving werden naderhand duizenden botten opgegraven. Een rivier had de ongelukkig omgekomen dieren meegesleurd, zegt Brusatte.
Langs de oostkust van de VS loopt een 1500 km. lange reeks van rif-bekkens. Vanaf Canada tot North Carolina. Ze liggen op de breuklijn waarlangs Pangea scheurde. Er ontstaan grote meren die weer uitdrogen door de hitte. Dit herhaalt zich vele malen. Vissen, dino's en Pseudosuchiers komen om in duizenden meters zandsteen, kleisteen en andere afzettingen. Hetzelfde gebeurde in Brazilië, Portugal en Marokko. Allemaal gelegen op dezelfde breuklijn. De temperatuur op aarde stijgt 3 à 4 graden. De megamoessons worden nog extremer. Van de plantensoorten sterft 95% uit. En daarmee amfibieën, dino's en zoogdierverwanten. Overal worden sporen van deze dieren gevonden in de afzettingen. Maar nadat de lavastromen veel wegvaagden gaan de sporen van veel nieuw ontstane dino's overheersen.
Grotere dino's gaan overheersen temidden van nieuwe soorten varens en Ginkgobomen. Van de Pseudosuchiers overleven alleen enkele primitieve krokodillen.
Er bestaat geen verklaring voor de overwinning door de sauriërs over al het andere dierlijke leven in het Trias.
En voorouders van de nieuwe klassen en orden van Sauriërs zijn onbekend.
Op het Schotse eiland Skye beginnen de gigantische Sauropoden op te duiken. Lange linten van kuilen in een patroon van link-rechts blijken de sporen van deze dieren. Samen met die van zoogdieren die er uitzien als bevers.
Sporen van Brontosaurus, Brachiosaurus en Diplodocus.
In het begin van het Trias (of einde-Perm) ontstaan de Lepidosauriers. Het zijn diapsiden, zoals alle reptielen. Er zijn echter alleen fragmenten en schedels gevonden. Tot de Lepidosauriers behoren hagedissen, brughagedissen en slangen. Over hun ontstaan is weinig of niets met zekerheid te zeggen.
In het Trias leven eveneens de Thecodonten. Dit zijn ook diapside dieren maar dan met tanden die met een wortel in de kaak zijn geplaatst. Dit komt voor bij krokodillen, zoogdieren en mensen.
Uit de Thecodonten zouden de Dinosauriërs, de Pterosauriers, de Ichtyosauriers en de krokodillen zijn ontstaan.
Een goed v.b. van een Thecodont is Thrinaxodon, een Dicynodont.
https://upload.wikimedia.org/wikipedia/commons/thumb/e/e2/Iziko_
Thrinaxodon_fossil.JPG/260px-Iziko_Thrinaxodon_fossil.JPG
Dit dier wordt beschouwd als een overgangsvorm naar de Therapsiden. Het vertoont allerlei geavanceerde elementen op weg naar de zoogdieren:
- gedifferentieerd gebit met kiezen, snijtanden en hoektanden
- kiezen multicuspidaat
- een snelle renner op vooral de achterpoten
- een secundair benig verhemelte
- kleine kanaaltjes in het neusbeen. Daarin liepen zenuwen en bloedvaatjes.
Dan ook snorharen. En indien snorharen dan waarschijnlijk een vacht.
- in de vacht zweetklieren en geurklieren?
- geen buikribben. Daarom een middenrif voor nog betere ademhaling.
Daarom ook warmbloedig (endotherm)??
- 7 halswervels aanwezig net als bij alle zoogdieren.
Bedenk hierbij wel dat de Dicynodonten synapside dieren zijn, net als de zoogdieren.
Alle Dinosauriërs zijn diapside dieren.
Weer zien we dat er toe geredeneerd wordt via allerlei onbetrouwbare tussenstappen naar het "zoogdier". Maar waarom zijn juist deze dieren dan op weg naar de ondergang??
Het Karoo-bekken is een heel unieke vindplaats. Hier werden in totaal wel 800 miljoen fossielen opgedolven van gewervelde dieren. Nog steeds is de beenderenvoorraad niet uitgeput. De zand- en kleisteenlagen zijn wel ruim 6000 m. dik.
De oudste Dino zou Staurikosaurus geweest zijn.
https://upload.wikimedia.org/wikipedia/commons/thumb/3/3d/Staurikosaurus_skeletal.jpg/260px-Staurikosaurus_skeletal.jpg
Er is maar 1 exemplaar van dit skelet gevonden in Brazilië. Het was een carnivoor van ruim 2m. lang en 20 kg. zwaar. Van dit exemplaar waren alleen vele wervels, de onderkaak, de bekkengordel en de achterpoten aanwezig. Het acetabulum had een verdikte lip aan de bovenrand van de gewrichtskom en een kleine opening in de kom. De mogelijke ontwikkeling naar een 2-benige, snelle gang met kleinere voorpoten (die niet gevonden zijn!) moet de start naar evolutie van de Dino's zijn.
Tegenwoordig rekenen de geleerden dit dier tot de fam. van Herrerasauridae. Deze fam. behoort dan tot de meest basale Therapoden of de stamgroep van de daar min of meer mee overeenkomende Saurischia. De moderne vogels worden eveneens tot de Therapoden of de Saurischia gerekend.
Onderweg naar "vogels" ?
Longisquama, ong. 12 cm. lang. Ze hadden veervormige schubben in de hals of op de rug. Nuttig voor gebruik als valscherm? Longisquama zoals gevonden als fossiel in steen
Longisquama zoals gevonden als fossiel in steen 
Gefantaseerde plaatsing van de "schubben". https://upload.wikimedia.org/wikipedia/commons/thumb/d/d1
/Longisquama_BW.jpg/220px-Longisquama_BW.jpg
Dit fossiel speelde een grote rol in de discussie of vogels nu wel of niet afstammen van Dinosauriërs. Sommige onderzoekerswaren van mening dat de "veren" niet anders waren dan toevallig aanwezige geveerde varenachtige bladeren.
Tegenwoordig wordt Longisquama als mogelijke eerste stap op weg naar de vogels niet meer serieus genomen. Dan komt misschien Podopteryx (Sharovipteryx) daarvoor eerder in aanmerking ? Maar hier is een vlieghuid gespannen tussen de staart en de achterpoten. Nuttig voort zweven. Voorouder van de Pterosauriers?
https://upload.wikimedia.org/wikipedia/commons/thumb/3/32/Sharovipteryx_mirabilis_fossil.JPG/250px-Sharovipteryx_mirabilis_fossil.JPG
Protoavis. 225 milj. jaar oude fragmenten van fossielen die vogelachtig waren. Dat is 75 miljoen jaar eerder dan de beroemde Archaeopteryx. Archaeopteryx moet daarom een doodlopende tak geweest zijn.
https://upload.wikimedia.org/wikipedia/commons/thumb/1/11/ProtoavisDrawing.jpg/266px-ProtoavisDrawing.jpg
Sommige wetenschappers beschouwen Protoavis als een samenraapsel van beenderen van verschillende diersoorten. De botten vormden geen skelet maar waren op een hoopje samengedreven. Allerlei geavanceerdere vogelachtige kenmerken werden aan Protoavis toegeschreven, zoals minder tanden, een betere vleugelconstructie, een furcula (sleutelbeen) en een sternum (borstbeen) met een grotere kiel. Daarenboven grote borst- en schouderspieren samen met punten voor veeraanhechtingen. In de botten bevonden zich ook pneumatische holtes voor een betere ademhaling.
Allemaal wensen voor de gedachte dat vogels al hun oorsprong hadden in het Trias van sauriërachtige dieren.
Pneumatische holtes in de beenderen kwamen voor bij vroege krokodillen en vele sauriërs. Ze zouden kunnen dienen als koelorganen en voor een betere ademhaling.
Nog een probleem: bij kippen en andere vogels bevatten de vleugels de vingers nr. 2, 3 en 4.
Bij de Dinosauriërs vindt er bij diverse groepen reductie plaats van de vingers door verlies van eerst nr. 5, vervolgens nr. 4 en tenslotte nr. 3. Dus blijven over de nrs. 1 en 2.
Bij Terrestrisuchus is de 1e vinger gereduceerd.
Aquatische reptielen.:
De oorsprong van deze dieren is in nevelen gehuld.
Ontstaan via Hemiprotosuchus? Een Crocodylimorph.
In het 'Berlin Ichthyosaur State Park, gelegen in de Sierra Nevada (VS) zijn vele fossielen van Shonisaurus gevonden. Ze waren ong. 15 m. lang. Er werden ong. 37 exemplaren bij elkaar gevonden. Ze waren levendbarend. De plaats van de fossielen doet denken aan gestrande walvisachtigen op huidige stranden.
https://upload.wikimedia.org/wikipedia/commons/thumb/9/95/Shonisaurus_BW_2.jpg/266px-Shonisaurus_BW_2.jpg
An het einde van het Trias sterven Aetosauriers, de Thecodonten en verreweg de meeste Therapsiden uit.
Maar de 'primitieve' Lesothosauriërs blijven voortbestaan tot het einde van het Krijt. Let op de lange achterpoten en de korte voorpoten. Ze hadden geen zijwaarts uitslaande poten, wel holle beenderen.
https://upload.wikimedia.org/wikipedia/commons/thumb/a/a1/Lesothosaurus_diagnosticus.png/266px-Lesothosaurus_diagnosticus.png
Gedurende de Jura.
"De zoogdieren spelen 140 milj. jr. een nietige bijrol".
Vanwege de veroveringen door de grote en kleine Dino's.
Dat is een hoogst vreemde redenering omdat de laatste tijd toch wel het besef is doorgedrongen dat aan het reuzenbestaan evenveel nadelen als voordelen kleven. En als zoogdieren uitgerust zijn met eierstokken, eileiders, baarmoeder, placenta melklieren een goed haarkleed kunnen ze vele malen beter overleven in bijna elk klimaat dan Dinosauriërs.
Stegosauriers en Ankylosauriers kunnen afgeleid zijn van de Lesothosauriërs of Scelidosauriers.

een Stegosaurier.

een Ankylosaurier.

een Scelidosaurier.
Een groot geslacht van wel 17 soorten Dino's is dat van de Cetiosauriers.
Cetiosauriers waren grote (16 m. lange) plantenetende Sauropoden. Met gestrekte, niet erg lange nekken en vrij kleine wervels. Ze hadden zich verspreid in alle windrichtingen. Sommige exemplaren droegen een zware knots aan de staart. Een goed verdedigingsmiddel voor een grote planteneter. In de groeve van Dashampu (in Sichuan, China) werden een tiental skeletten van Shunosauriers gevonden. Dit zijn verwanten van de Cetiosauriers; grote planteneters.
De kudde zou moeten zijn meegesleept naar hun ondergang in de groeve. Maar wel samen met Carnosauriers, Coelosauriers, Ornithosauriers, Pterosauriers, Lesothosauriers, Yandusauriers, vissen en Labyrentodonten (grote amfibieën). Deze alle vermengd met fossiele hoge boomstammen.
De Tendaguru Formatie in Tanzania vertoont een vergelijkbaar beeld. Zoet- en zoutwaterafzettingen wisselen elkaar daar af en bevatten de skeletten van Sauropoden (o.a. Brachiosaurus, Barossaurus, Dricraeosaurus, Tornieria), Stegosauriers, Ornithischiers, Theropoden en Pterosauriers.
Dan nog de beroemde Morrison Formatie in de VS (1 miljoen km2 groot) met Brontosauriers, Diplodocus en Camarosaurus.
Vele sauriërs leefden langs rivieren, meren en zeeën.
Samen met duizenden andere fossielen. Bedenk daarbij wel dat er vele grote reuzen waren, tot 27 m. lang onder de bovengenoemde sauriërs en vele, vele andere; grote en kleine. Zoek ze allemaal maar op in Wikipedia!
Tussen de Bighorn Mountains en Yellowstone Park zitten de rode en grijze rotsen vol met dino's. O.a. Brontosaurus, Brachiosaurus en Allosaurus (de grote eters).
Op de grens van Utah en Colorado 120 sauriërs en tienduizenden botten van Allosaurus.
In Oklahoma wordt een beenderlaag aangetroffen bij een wegaanleg door werkloze arbeiders
Een aanvaardbaar verhaal over de oorzaken en de bewaring van al deze fossielen wordt niet geleverd.
Evenmin als de stamvaders van al de genoemde groepen. Suggesties genoeg over de cladistische plek van vele Dino's en andere sauriërs, maar geen bewijzen.
Honderden skeletten van sauriërs aanwezig in:
- Dinosaur National Monument in Utah
- de Cleveland-Lloyd-groeve in Utah
- Dry-Mesa-groeve en Garden Park in Colorado
- Bone Cabin en Howe Querry in Wyoming.
De botten van verschillende soorten liggen vaak opeengestapeld: 20 skeletten en 4000 botten. Dankzij elkaar opvolgende catastrofes. Een heel ecosysteem van Diplodocus, Barosaurus, Allosaurus en Camptosaurus. Zijn de dieren vast gebleven in de modder na een overstroming?
Of was er een natuurlijke valstrik voor de rovers onder hen? Of was er een Allosauriër of grote planteneter vastgeraakt in de modder? Het roofdier raakte daarop zelf verstrikt in de modder. Dit dier trok weer andere hongerige Allosauriërs aan etc. Losgeraakte botten spoelden naderhand weg. Veel beenderen zijn gebroken. Aldus een zeer onwaarschijnlijk verhaal!
In de Dry-Mea-groeve wel 100 ton aan botten opgegraven. Waaronder Torvosaurus, Supersaurus van 40 m., Ultrasaurus van 25 m., Diplodocus van 30 m., Seismosaurus 32 m.

Seismosaurus.
In de kustgebieden van deze omgeving leefden Plesiosauriers en Ichtyosauriers.Hoe deze dieren hier bewaard bleven??
In Midden-Europa werden hoge kalsteenformaties afgezet, b.v. in Solnhofen. Met hierin fossielen van Sauropoden samen met kleine vliegende reptielen (Pterosauriers) waaronder de beroemde Rhamphorynchus. Verwant aan de Pterodactyloidea. Ze waren verspreid over heel Europa. Er zijn wel 100 complete skeletten van deze dieren gevonden. Soms werden in de 1 mm. dikke structuur van de huid nog blijken van bloedvaten en spierweefsel aangetroffen. Over de oorsprong van deze diergroepen tast men volledig in het duister.
https://upload.wikimedia.org/wikipedia/commons/thumb/1/11/Rhamphorhynchus_longicaudus.jpg/260px-Rhamphorhynchus_longicaudus.jpg
In de groeven van Solnhofen werd ook het wereldberoemde skelet van Archaeopteryx ontdekt. Dit zou de overgang tussen sauriër en vogel markeren, meenden de geleerden decennialang. Hier is men intussen wel van teruggekomen. De "vogel" had geen luchtholtes in de beenderen, geen borstbeen en zwakke vliegspieren. Veel later ontdekte gevederde sauriërs hebben Archaeopteryx van zijn bevoorrechte plaats verdrongen. Van Archaeopteryx is geen enkele voorvader bekend.
Vele fossielen vertonen een 'krampachtige' achterover gewrongen positie. Er is zelfs gesuggereerd dat vele dieren tezamen door stormen in het water geblazen zijn. Zo is voor elk probleem een "goed" verhaal.
5.4 Gedurende het Krijt.
Het klimaat verandert tijdens het Krijt geleidelijk van een broeikasperiode naar kouder en droger. Samengaand met een grote stijging van de zeespiegel en daarna een sterke daling. Zuid-Amerika en Afrika wijken uiteen. Er zijn geen poolgebieden met ijskappen. Tenslotte betekende dit het uitsterven van de Dino's.
Europa was waarschijnlijk in grote eilandengroepen verdeeld; Noord-Amerika was in tweeën gedeeld door de Binnenlandse Krijtzee. Noord-Afrika, Zuid-Amerika en India stonden grotendeels onder water en India verplaatste zich naar het noorden. Alle gebieden waren heel divers betreffende aanwezige dieren en planten. Noord-Amerika zag er misschien als volgt uit:
Op de zeebodems ontstaan enorme dikke, zwarte modderlagen. Mede dankzij zuurstofgebrek.
Daarin vinden Dino's, andere reuzewaterdieren zoals Plesiosauriers en schildpadden, samen met zoogdieren de dood. Van de laatste waren het over het algemeen kleine exemplaren. Dus werden ze overheerst door de grote sauriërs?? Een merkwaardige gedachte!
In de modderlaag ook vele Angiospermen (bloemplanten).
De Titanosauriërs (b.v. Argentinosaurus) verschijnen samen met de Ornithischiers.
Ankylosauriers (met hun grote beenplaten) bewonen het land i.p.v. de Stegosauriers.
De Ornithischier Iguanodon verovert het hele aardrijk. Hieruit zouden de Hadrosauriers (de eendesnalvelsauriers) zijn voortgekomen. Het zijn allemaal planteneters. Maar ook eters van zaden, noten, insecten, vissen en schelpdieren komen voor.
Iguanodon is ong. 10 m. lang. Met 5 tenen aan de voorpoot (met de duim als een grote stekel) en 4 tenen aan de achterpoot. Ze zouden moeten zijn ontstaan uit Camptosaurierachtige voorouders.
De Iguanodonten leven in heel Europa tot in de Kaukasus en verspreiden zich ook in de VS en Mexico. Vandaar steken ze over naar Zuid-Amerika en Afrika.
Er worden 30 soorten onderscheiden.
De eerste, wel 30 complete skeletten, worden in België (Bernissart) ontdekt in een heuvelrug.
Iguanodon is de eerste Dino die als volledig skelet wordt beschreven.
De Sauropoden zijn vooral de grote planteneters. Met lange nekken en kleine hersenen.
Eind-Trias ontstaan de kleine Protosauropoden. Gedurende de Jura verspreiden ze zich over de hele aarde. In het Krijt verdwijnen de Sauropoden van het noordelijk halfrond (Laurazië) en vinden we de grote jongens (Brontosaurus, Brachiosaurus en Diplodocus) op Gondwanaland.
Op het Isle of Sky in Schotland worden rijen sporen van grote Sauropoden, samen met zwemmende zoogdieren, zo groot als bevers, aangetroffen.
Het gewicht van een zware sauropode kan worden vastgesteld door de botten te wegen en door m.b.v. fotogrammetrie en animatiesoftware de omvang van het dier te bepalen. Zo komen we aan gewichten tot wel 50 ton. Vgl. met een olifant die ong. 10 ton weegt.
Hoe konden de Sauropoden een dergelijk gewicht verkrijgen?
1. door met behulp van een lange nek veel voedsel te verzamelen
2. door een snelle groei. In 30 tot 40 jaar volwassen
3. door bezit van een zeer efficiënte long.
4. door de lichter bouw vanwege de holle beenderen
5. door de hogere lichaamstemperatuur een veel grotere warmteafgifte.
De zuurstofvoorziening is te vergelijken met die van vogels. Dankzij het bezit van extra luchtholtes, ook in de botten, stroomt de in- en uitademingslucht in één richting. Dit heeft een veel betere zuurstofopname tot gevolg.
De Theropoden (vleeseters) hadden zo'n longsysteem, de Ornithischiers (planteneters) niet.
Een bijzondere Therapode was Deinonychus. Dit dier had gigantische klauwen aan beide poten. Misschien was het een groepsjager. In ieder geval was het een vervaarlijke rover. Het skelet lijkt op dat van een Archaeopteryx.
Veel fossielen werden gevonden in de Sahara (Niger) en in Marokko.
Honderd ton aan botten werd verzameld.
Langs zeekusten met mangrovebossen en rivierdelta's leefden schelpdieren, vissen. samen met vele sauriërs.
Intussen waren er al meer geavanceerde vogels ontstaan:
Ambiortus. Het skelet vertoont een gekield borstbeen, een verlengd ravenbeksuitsteeksel en vergroeide handbeentjes.
Ambiortus. Gereconstrueerd skelet naast geïdealiseerde afbeelding .
bron Bing.com.
Enaliornis of Pelagornis. Een tot de zwemmers behorend vogelgeslacht; lijkend op een flamingo?
Met een vleugelspanwijdte van 6 à 7 m. Ontstaan tijdens Mioceen? (bron Wikipedia).
Yanornis. Ontstaan in vroeg-Krijt.
Gevonden fossiel van Yanornis met geïdealiseerd "portret". (bron Wing. com).
Andere v.b. van dergelijke dieren zijn gevonden in Spanje, China en in Australië.
Vele van deze fossielen worden gerekend tot de Ornithomorpha.
De voorouders van deze dieren kan van alles geweest zijn:
- een Dino
- een Notosuchier of ander "krokodil"
- een Thecodont.
Notosuchiers waren land bewonende krokodillen met zoogdiertanden. Ze leken op vroegere zoogdierachtige reptielen. Hun afkomst is onzeker.
Ichtyosauriers en Plesiosauriers sterven uit in Midden-Krijt. Waardoor??
De Coelurosauriers waren in Laurazië wijd verspreid maar in Gondwanaland uitgestorven.
Tot deze groep behoort ook Tyrannosaurus rex en de orde Maniraptoriformes. Deze dieren bezaten protoveren. En waren warmbloedig; denkt men.
In Brits Columbia werden sporen van diverse sauriërs gevonden samen met die van kustvogels.
Vele skeletten van Protoceratops werden gevonden in Mongolië. Daarbij vele nesten met eieren van verschillende soorten dino's. In dit gebied ook skeletten van Oviraptor, Velociraptor. en Deinonychus (een vervaarlijke rover, met enorme klauwen, rennend op 2 achterpoten).
Ook van Deinonychus bestaan geïdealiseerde afbeeldingen met gevederde voorpoten. Voor Deinonychus geldt dat hij een vogelachtig lijf heeft en vergelijkbare lichaamsverhoudingen.
Belangrijke vraag bij bovengenoemde drie diersoorten is of ze wel of niet konden vliegen.
Samen met Velociraptor en Microraptor wordt Deinonychus ondergebracht bij de Dromaeosauridae.
De Dromaeosauridae hadden bevederde poten en waren daarmee, denkt men, nauw aan de vogels verwant. Misschien waren de voorouders echte vogels die het vliegvermogen hadden verloren.
Opm.: Zo kan elke redenering elke theorie ondersteunen!
Zijn de Coelurosauriers de stamvader van Oviraptor? Ze hebben een gedegenereerd gebit en een snavel aan de voorkant van hun kop.
Oviraptor behoort tot de Maniraptora. De nieuwste definitie van Maniraptora zegt: De groep die de huismus (Passer domesticus) en alle soorten die dichter bij Passer staan dan bij Ornithomimus omvat.

Vindplaatsen van fossielen van
Ornithomimus.
Een massagraf van tienduizend Maiasaura bleek aanwezig in Montana (op de grens met Wyoming). Samen met duizenden stukken bot van Deinonychus. Daar zijn de Rocky Mountains ontstaan in Laat-Krijt, tijdens de Laramide-orogenese. Het massagraf is overdekt met vulkanische as.
Was er sprake van enorme kudden van deze dieren? Jong en oud waren tezamen omgekomen. Er waren grote broedkolonies.
De Maiasauridae behoren tot de Hadrosauriers, de eendesnalvelsauriers. Maar de oorsprong van deze diergroep is een mysterie.
Van de 6 soorten Ceratopsiers (o.a. Centrosaurus, ong. 400 dieren) kwamen honderden grote dieren om bij het oversteken van een sterk stromende rivier. De beenderen lijken kapot getrapt door rovers. Ze zijn bewaard in massagraven in Alberta (Canada)
Volgens het evolutieverhaal stamden de Triceratopsiden af van kleine rondrennende bladtandige beestjes uit de vroeg-Jura. Zoals Heterodontosaurus en Lesothosaurus.
Ze liepen liever op 4 poten en ontwikkelden diverse vormen van horens en kammen op hun koppen. De eerste Ceratopsiden waren zo groot als een hond. Ook dit lijkt weer veel op het "paardenverhaal" in latere tijden. Hun kaken en tanden veranderden zodanig dat ze grote hoeveelheden bladen konden vermalen Een vlijmscherpe snavel hielp bij het afsnijden van takken en bladeren.
Van de Ceratopinae werden rond Hell Creek duizenden skeletten gevonden samen met vele andere sauriërs. En vele massagraven met "kudden" van wel 1000 dieren.
Tussen al deze sauriërs en vele grote Pterosauriers werden vele soorten buideldieren, (placentale)zoogdieren, haaien, beenvissen, steuren en roggen aangetroffen. Zelfs enkele uitgestorven vogels. Ze worden gerekend tot de grote groep van Enantiornithes. Wil je weten welke dieren allemaal werden gevonden in Hell Creek, zoek maar op in Wikipedia.
De Enantiornithes bevat de Maniraptora. Samen met Deinonychus e.a. vormen zij de Eumaniraptera.
De uitgestorven vogels die eveneens tot de Enantiornithes behoren zijn b.v.
De kiel op het borstbeen lijkt te ontbreken. Konden deze vogels vliegen?
De vogel leefde als een ijsvogel. Dus hij kon goed vliegen!
Het zal uit bovenstaande afbeeldingen duidelijk zijn dat het hier om vogels gaat. Om de overgang vanaf Maniroptera aannemelijk te maken wordt er wel gewezen op 'primitieve kenmerken' zoals te doen gebruikelijk is in de evolutietheorie. Zo heeft Sinornis tanden in de onder- en bovenkaak.
Maar deze dieren hebben een echt verenkleed en konden goed vliegen i.t.t. al de andere sauriërs met een "verenkleed".
Intussen is er nog steeds veel discussie onder geleerden over de ontstaansroute van de echte vogels.
In het vroeg-Krijt, toen deze vogels rondvlogen, groeiden er (de eerste?) bloemetjes in Azië. Zestig miljoen jaar later ook vele soorten bomen en struiken, samen met de grassen. Vele dieren aten zaden van Angiospermen.
Goiàs in Brazilië bevat een 300 m. dikke gesteentelaag. Uit het laat-Krijt. Hier worden de enorme Dino's opgedolven van 25 m. lang of meer.
Dat de grootte van een bepaald soort organisme geen belangrijk kenmerk is blijkt bij de mens.
Reuzengroei bij mensen leidt tot een lengte van 225 of zelfs 250 à 260 cm.
De kleinste mensen daarentegen zijn vaak ong. 50 à 60 cm. lang.
Dus een vijfvoudige lengtetoename is geen enkel probleem om toch een volwaardig menselijk leven te kunnen leiden met al de basale organen die daarvoor noodzakelijk zijn.
Toegepast op de Dino's betekent dit dat een Dino van 5 m. lang gemakkelijk uitgegroeid kan zijn tot een exemplaar van 25 m. Dit onder gunstige omstandigheden of bij veel concurrentie met sterkere dieren.
Zonder dat er fundamentele veranderingen in de bouw vereist waren!
5.6 Evolutie van de Dinosauriers.
Hoe verliep nu in grote lijnen de evolutie van de intussen gigantisch grote groep Dino's?
Het is van groot belang om je te realiseren dat een natuurwetenschappelijke benadering van deze vraag onmogelijk is. De fossielen zijn dood of levenloos. Waarneming en experiment zijn daarom principieel onmogelijk. Alle gevonden exemplaren op een rijtje zetten en letten op overeenkomsten is een aardige onderneming maar lost weinig tot niets op. Vergelijk dit maar met een verzameling van alle auto's of vliegtuigen. Leuk om naar te kijken maar geen enkele reden om te denken dat deze vervoers- mogelijkheden uit elkaar ontstaan zijn in de hoofden van de ontwerpers!
Op basis van de evolutieleer was de eerste sauriër een kleine 2-voetige vleeseter die de voorouder werd van de Ornithischiers, de sauriërs met een vogelachtig bekken. D.w.z. een bekkengordel met het ischium (het schaambeen) naar achteren gericht. Bij de Saurischia is het schaambeen naar voren gericht.
Beide hoofdgroepen samen waren de Dinosauriformes.
Ze zijn in het Trias ontstaan, 230 milj. jr. geleden. Tegen het einde van het Trias sterven de andere grote dieren op het land uit. Waardoor ?? De Dino's worden nu 140 milj. jr. de heersers op aarde.
Een hoogst merkwaardig idee overigens dat ontleend is aan de mensenwereld: hoe groter, des te sterker. In de dierenwereld speelt grootte in dit opzicht geen rol. Kleine en zeer kleine dieren komen in alle leefgebieden overvloedig voor samen met grotere exemplaren.
De als eerste beschreven sauriër was Iguanodon uit België.
Voor de indeling van alle Dino's wordt tegenwoordig gebruik gemaakt van het cladistische systeem.
Bij een clade neemt men niet de voorouder als uitgangspunt maar de verwantschap tussen afstammelingen van de voorouder. De computer berekent de graad van verwantschap op basis van een aantal overeenkomsten. Essentiele voorvraag bij elk ingedeeld organisme is hierbij natuurlijk welk(e) kenmerk(en) belangrijk genoeg zijn om mee te dingen als criterium voor plaatsing in de rangorde die aangebracht wordt.
Aangezien de voorouder van elke groep (clade) binnen de Sauropsida (alle reptielen) altijd onbekend is is het hele systeem een abstract geheel van "computerorganismen". Je kunt ook zeggen: het hele systeem valt als een kaartenhuis in elkaar als je er tegen stoot. Dit mag al overtuigend duidelijk zijn als je het samenvattende plaatje ziet van de Dino's:
Het musje voelt zich helemaal thuis tussen de Theropoden, de Stegosauriers en de Ceratopsiers.
De vogels zijn de overgebleven reptielen na alle rampen die over de aarde gingen.
Oftewel: de vogel is een reptiel voor de "gelovigen".
De definitie van de Sauropsida luidt: alle nakomelingen van de gemeenschappelijke voorouder van Iguanodon (een Saurischier) en Megalosaurus (een Ornithischier).
Sinds 1993 is de definitie verankerd op Triceratops horridus en Passer domesticus.
Dit gebeurt op basis van synapomorfieën omdat een bepalende eigenschap van een voorouder onbekend is. Het lijkt waarschijnlijk dat die "eigenschap" een combinatie is van:
- een grijpende hand
- een half-opponeerbare duim
- een verkorte 4e en 5e vinger.
Deze "eigenschap" biedt aardig wat mogelijkheden om het evolutieverhaal verder in te vullen totdat we arriveren bij ons musje!
Vogels stammen dus af van d Saurischiers. Bij deze organismen is het schaambeen naar voren gericht. Merkwaardigerwijs is bij sommige Saurischiers het schaambeen naar achteren gericht. Dit zou ontstaan zijn doordat ze planten gingen eten en het vergrote darmkanaal dit been naar achteren duwde.
Van de Ornitischiers is de herkomst volslagen onbekend.
Genoemd worden:
- Pisanosaurus; één skelet gevonden. Met een geavanceerde kop (??) en een primitieve romp (??). Was het een chimaer? Was er sprake van convergente evolutie ? Of een basaal lid (??) van de Dinosauriformes?
- Eocursor.https://upload.wikimedia.org/wikipedia/commons/thumb/7/71/Eocursor_BW.jpg/266px-Eocursor_BW.jpg
Een "vogelachtige" reconstructie. Slechts één skelet.
In Liaoning (China) werden en worden duizenden fossielen opgedolven van meer dan 20 soorten. Deze zijn hier terechtgekomen dankzij vulkaanuitbarstingen. Het betreft vooral kleine dino's met kransen van veerachtig dons. Voorbeelden zijn:
- Sinosauropteryx
https://upload.wikimedia.org/wikipedia/commons/thumb/b/b8/Sinosauropteryx_GMV_2124.jpg/270px-Sinosauropteryx_GMV_2124.jpg
Was het dier bekleed met veren of haren? Wat is haarachtig dons?
Was het dier warmbloedig? Had het luchtzakken of ademde het m.b.v. een middenrif?
- Caudipteryx
Het dier had veren aan de voorpoten en staart. De veren waren symmetrisch, dus niet geschikt om mee te vliegen. Een veer was een schacht met zijtakjes.
- Protarchaeopteryx
Het dier had symmetrische veren aan de staart.
- Microraptor.
Het dier had een enorm lange staart. Het had veren aan de voor- en achterpoten en aan het uiteinde van de staart. Ook hier bestonden de veren uit schacht en baarden.
Volgens Brusatte zijn de vogels Theropoden, waartoe ook de reusachtige vleesetende dino's behoren. De bovengenoemde dieren hebben veren, een vorkbeen, drievingerige voorpoten en honderden andere aspecten van het skelet die overeenkomen met die van de vogels. Dat samen moet wel betekenen dat vogels afstammen van sauriërs. En: ze gedroegen zich zonder meer als moderne vogels. Hoe de schrijver dat toch heeft waargenomen!?
Als we dit soort 'hoogvliegerige' uitspraken wat kritischer analyseren komen we tot heel andere conclusies.
Om te beginnen bij de vogelveer. Er is een holle schacht met baarden die weer bezet zijn met baardjes die met behulp van duizenden weerhaakjes in elkaar grijpen. Door dit ingenieuze systeem kan een vleugel vliegen mogelijk maken. Maar er zijn diverse soorten veren: slagpennen, dekveren en donsveren. Elke soort veer moet dankzij genetische factoren op precies de juiste plaats worden ingeplant en tot de juiste lengte uitgroeien. Voor elke soort kenmerkend. Daarbij komt nog het prachtige kleurenpatroon van alle veren samen die een vogelsoort herkenbaar maken en essentieel is voor de paarvorming. Of kan als schutkleur of dreigmiddel wordt gebruikt.
Vervolgens: het skelet met de de zeer compacte bouw van wervelkolom, bekkengordel, vergroeide vingerkootjes (de nrs. 2,3 en 4), het vorkbeen, het langwerpige smalle schouderblad en het borstbeen met een grote kiel. 350px-Squelette_oiseau.JPG (350×466) (wikimedia.org)
Nog veel ingewikkelder wordt het als we de manier van ademhalen gaan bestuderen. De ingeademde lucht stroomt eerst door de longen en vervolgens naar 9 luchtholtes verdeeld over het lichaam.
Eerste fase: de lucht stroomt naar de achterste 4 luchtzakken en richting de longen
Tweede fase: de zuurstofrijke lucht stroomt door de longen naar de 5 voorste luchtzakken.
Derde fase: de zuurstofarme lucht wordt uit de voorste luchtzakken naar buiten geperst.
Bij de daaropvolgende inademing stroomt er verse lucht naar de achterste luchtkamers en tegelijkertijd door de longen. Bij het uitademen wordt de lucht vanuit de voorste luchtkamers naar buiten geperst.
Bij dit in- en uitademen zijn spieren die borstbeen en ribben verwijden actief.
De holle beenderen vormen enige opslagruimte voor zuurstofrijke lucht.
De holle beenderen bij vele soorten dino's en het misschien warmbloedig zijn van b.v. de raptors kan geen enkel bewijs vormen voor het ontstaan van vogels uit sauriërs.
Als een dier(tje) gaat vliegen zal het duidelijk zijn dat de hersenen hierop helemaal berekend moeten zijn. Hoe stijg je op? Hoe regel je heel perfect de vleugelslag en positie van de staart bij koerswijziging en landen op een tak, nest of op het land/water? Hoe leer je vliegen? Hoe leer je je veren verzorgen?
Een vogel moet 2 keer per jaar ruien. Dit wordt nauwkeurig geregeld: snel in één keer of geleidelijk over langere tijd. Vaak in prachtkleed in voortplantingstijd!
Als bovenstaande allemaal mogelijk gemaakt is moeten zowel bloedsomloop, zenuwstelsel, voortplantingsstelsel en zintuigstelsel enz. er passend bij worden aangelegd. Er steeds bij bedenken dat voor elke wijziging in bouw van een onderdeel een ontwikkelingsproces vanuit de ene eicel noodzakelijk is.
Vereist voor het ontstaan van een vogel is daarom een genenstructuur in het DNA van duizenden nieuwe perfect georganiseerde genen. En daarbij aansluitend de verschillende RNA's en mikrosomen. Dat betekent duizenden nieuwe eiwitten die allemaal hun plaats en functie gewezen wordt door genen en vele andere moleculen die dit weer aansturen.
Het boek van Stuart Burgess 'Ontwikkeling of ontwerp' (uitg. Medema) besteedt uitvoerig aandacht aan de vogels met hun structuur van de vleugels, de structuur van de veren, de migratie van veel vogelsoorten, de gevonden fossielen van vogels en hun stofwisseling. Vervolgens bespreekt hij de ongelofelijk ingewikkelde bouw van de pauwenveer en de zang van zangvogels. De nodige tussenstappen voor de evolutie van een pauwenstaart zijn absoluut onmogelijk. De schrijver noemt wel 9 met elkaar samenhangende structuurfactoren. Daarvan zijn de kleurpatronen dankzij de dunne-laaginterfferentie samen met de mathematische structuren binnen het 'oog' wel de meest wonderbaarlijke.
Ondanks alle prachtige en wetenschappelijk ogende cladistische schema's mogen we wel concluderen dat ze berusten op zeer twijfelachtige aannames en constructies. Het betreft veelal toeredeneren naar vooraf bepaalde ontwikkelingslijnen die moeten harmoniëren met de ouderdomsbepalingen.
Het is belangrijk om te bedenken.dat Pterosauriers (vliegende reptielen), Ichthyosauriers, Plesiosauriers en Mosasauriers (de laatst drie zwemmende dieren) geen sauriërs zijn.
Interessante vraag: waar kwamen al deze dieren vandaan?
Hoofdstuk 6
Naar een herziene aardgeschiedenis (geologie)?
In het boek van Jens Boeningk, Sabine Wodniok en Edvard Glücksman Biodiversity and Earth History wordt een prachtig overzicht geboden van alle geologische perioden en de bijbehorende diverse levensvormen. Bij de indeling van de organismen wordt uitgegaan van het cladistische systeem.
De drie gezamenlijke criteria hiervoor zijn:
- de structuren van DNA en RNA
- stofwisselingsprocessen
- morfologische overeenkomsten en verschillen.
Bij de bespreking en beoordeling daarvan ga ik eerst de complete evolutionaire indeling langs en vervolgens de verdeling van de groepen organismen tijdens de aardgeschiedenis. Bij dat laatste ga ik uit van de veronderstelling dat de periode Trias-Jura-Krijt een wellicht mogelijk uitgangspunt kan zijn voor de indeling van de aardgeschiedenis. De periode Krijt omdat het einde daarvan gemarkeerd wordt door een geweldige catastrofe waarbij er een einde kwam aan het Dinosauriertijdperk door de inslag van, zo meent men op goede gronden, een enorme meteoriet. Bovendien omdat tijdens de Krijtperiode alle groepen van het dieren- en plantenrijk reeds aanwezig waren.
Ik ga er vanuit dat Pangea bestond vanaf de schepping van de aarde.
Vanaf laat Devoon/Carboon begint Pangea door allerlei processen uit elkaar te gaan.
Tijdens het Krijt begint dit aardrijk sneller uiteen te vallen.
De supergroepen.
(de structuur van de cellen getekend naar het boek Biodiversity and Earth History).
6.1 Unikonta.
Hiertoe worden gerekend de Opisthokonta (de flagel achterwaarts gericht), de Amoebozoa (amoeben en slijmzwammen met schijnvoetjes) en de Apusozoa.
Tot de Opisthokonta behoren de Choanomonada (de sponzen) en alle overige "dieren" evenals de "schimmels" (de Holomycota).
De Apusozoa hebben 2 flagellen. Ze zijn dus bikont. Ze worden ook wel tot een andere supergroep van het moderne cladistische systeem gerekend, nl. de Rhizaria. Maar deze hebben allemaal amoeboïde cellen (dus met schijnvoetjes) Daartoe behoren Foraminifera en Radiolaria.
 Een Foraminifeer.
Een Foraminifeer.https://upload.wikimedia.org/wikipedia/commons/a/a6/Ammonia_tepida.jpg
De Choanomonada zijn daarentegen unikont.
Veel cellen van Metazoa (alle "dieren") zijn unikont. Denk b.v. aan hun zaadcellen die ook ophisthokont zijn.
Maar het schijnt zo te zijn dat de Unikonta afstammen van bikonte flagellata omdat er aan de basis van de flagel een tweede kinetosoom aanwezig is.
Cel van een Choanomonada.
1. flagel 2. krans van 20 à 40 microvilli 3. kern 4. endoplasmatisch reticulum 5. lorica.
De
lorica is wel of niet aanwezig. Ze bestaat uit
cellulose of
silicium. De cellen zijn soms vastgehecht aan een substraat.
De flagel doet water wegstromen van de flagel, via de microvilli. De microvilli vangen voedseldeeltjes op en transporteren die naar het cytoplasma.
Mitochondria zijn ook aanwezig.
Kraagcellen in een spons (Porifera) hebben eenzelfde bouw en werking.
De netelcellen van de holtedieren hebben een totaal andere bouw en functie.
De andere basale groepen van de Holozoa zijn 1-cellige parasieten.
De fylogenetische relatie binnen de diverse groepen van Unikonta is zeer onzeker. De Apusozoa zijn waarschijnlijk een parafyletische groep.
Moleculair fylogenetisch onderzoek biedt voorlopig weinig zekerheid voor het vinden van voorouders binnen het cladistisch systeem.
En: de indeling van alle organismen op grond van het aantal flagellen is erg onbetrouwbaar.
6.1.1 Holozoa.
De Holozoa omvatten zeer heterogene groepen zoals 1-celligen, sponzen, insecten, wormen, schimmels, vissen, zoogdieren en mensen.
De fylogenetische ontwikkelingslijnen binnen de Holozoa blijven nog steeds onduidelijk!
Vaak zijn er exoskeletten ontwikkeld; al ver voor het Cambrium. Tijdens de zgn. Cambrische explosie.
Binnen vele groepen vindt er parasitisme plaats.
Uitgezonderd de sponzen hebben de Holozoa 2 kiemlagen tijdens hun vroege ontwikkeling.
De organismen binnen hoofdgroep Bilateria zijn tweezijdig symmetrisch.
Alle Bilateria, uitgezonderde de Cnidaria, hebben 3 kiemlagen tijdens hun vroege ontwikkeling.
a. Choanomonada.
Eén- of meercellige organismen met 1 flagel. De flagel heeft 2 tubuli in het midden en 9 tubuli daar omheen. De flagel is omgeven met 20 à 40 microvilli die actine-filamenten (vgl. onze spieren) bezitten. De microvilli zorgen zorgen voor de voedselopname. Het voedsel wordt verteerd in een voedselvacuole en uitgescheiden door een uitscheidingsvacuole (vgl. het pantoffeldiertje).
De Choanoflagellaten zouden homoloog zijn aan de choanocyten (kraagcellen) van de sponzen.
Afstamming van beide groepen en van alle Holozoa blijft een discussiepunt.
Hoofdvraag: had de voorvader één of twee flagellen? Vaak is een kinetosoom aan de basis van de flagel aanwezig. Dus een gereduceerde flagel?
b. Porifera (sponzen).
Porifera hebben geen zenuwcellen of spierweefsel. Geen darmkanaal of voortplantingsklieren. Kraagcellen (met 20 à 40 filipodia) zorgen voor voedselopname. Kalk- of kiezelnaalden worden gevormd in de buitenste cellaag. Larven ontstaan bij kalksponzen door gastrulatie. Sponzen worden daarom wel beschouwd als model voor evolutie van de metazoa.
Voortplantingscellen kunnen uit al de 5 typen cellen ontstaan.
Als je een spons door een fijnmazig zeefje perst hergroeperen de cellen zich tot een nieuwe spons.
c. Placozoa (plakdiertjes).
De dieren hebben enkel ectoderm en endoderm (diploblast).
Slechts 1 soort binnen de eenvoudigste stam (rijk) van het dierenrijk met slechts 4 soorten cellen.
Geplaatst voor of na de Porifera of zelfs na de Cnidaria.
In 1883 ontdekt in een zeewateraquarium. Bleek later op diverse plaatsen aanwezig.
Niet symmetrisch. Geen zenuwstelsel.
Mogelijk ook aan de oorsprong van het dierenrijk?
https://upload.wikimedia.org/wikipedia/commons/thumb/c/c3/
Trichoplax_mic.jpg/266px-Trichoplax_mic.jpg
d. Cnidaria (koralen, poliepen, kwallen en zeeanemonen).
Ze zijn radiair symmetrisch en bezitten netelcellen op hun tentakels Ze hebben een netvormig zenuwstelsel met een zenuwring rond de mondopening. Geen spiercellen. Elke cel bezit zich samentrekkende vezeltjes.
Fossiele koralen bestaan vanaf het Ordovicium.
e. Ctenophora (ribkwallen).
https://www.bing.com/th?id=OIP.dcq7lVTRuHRMv8byy1ySpgHaFF&w=254&h=160&c=8&rs=1&qlt=90&pid=3.1&rm=2
Ze zijn niet-symmetrisch en hebben geen netelcellen. Ze vangen hun prooi m.b.v. colloblasten. Deze bevatten een opgerolde spiraal in de cel die naar buiten kan schieten en zich hecht aan een prooi.
Ze bezitten spierweefsel en hun ontwikkeling vertoont overeenkomst met de tripoblasten.
Ze bezitten lijsten van trilharen voor de voortbeweging.
Geen fossielen en de phylogenetische plaats is onduidelijk.
6.1.2 Protostomia
Ze worden beschouwd als een zustergroep van de Deuterostomia.
Bij de Protostomia ontstaat de mond uit de blastopore (de opening van de blastula).
Bij de Deuterostomia ontstaat de anus uit de blastopore.
Fylogenetische relaties binnen de Protostomia blijven onopgelost!
Anneliden (Spiralia) en Arthropoden vertonen overeenkomsten betreffende de gesegmenteerde lichaamsbouw, een touwladderzenuwstelsel en lengtespieren. Ook vergelijkbare extremiteiten?
Maar zijn de monddelen en gelede poten van de insecten homoloog aan de parapodia van de wormen?
Dezelfde vraag betreft de lichaamsholtes van beide groepen.
Moleculaire studies weerspreken de verwantschap.
a. Ecdysozoa.
Deze dieren bezitten een driedelig chitine-skelet. Ze hebben een door hormonen bestuurde ontwikkeling via vervellingen. Aangepaste flagellen hebben een sensorische functie en liggen in de cuticula. Ze hebben geen trochophoralarve.

bron Wikipedia.
Deze groep omvat rondwormen, geleedpotigen (insecten, spinnen, kreeften), duizendpoten e.a.
De spoelwormen of rondwormen (25000 soorten zijn beschreven maar omvat misschien wel 10.000.000 soorten) hebben 4 zenuwstrengen in de lengte (dorsaal, ventraal, lateraal) en een ring rond de pharynx.
Ze zijn niet gesegmenteerd. Ze hebben wel mannelijke of vrouwelijke voortplantingsorganen.
Tot de
Tardigrada behoort o.a. het
Beerdiertje. Duidelijk gesegmenteerd.
https://upload.wikimedia.org/wikipedia/commons/thumb/c/cd/SEM_image_of_Milnesium_tardigradum_in_active_state_-_journal.pone.0045682.g001-2.png/266px-SEM_image_of_Milnesium_tardigradum_in_active_state_-_journal.pone.0045682.g001-2.png
Het diertje kan leven onder de meest extreme denkbare omstandigheden: van +149 graden C tot
-272 graden C. Ze verdragen UV-licht en gammastraling. En dagenlang verblijf in de ruimte.
De Arthropoda zijn gesegmenteerd met een 2-delig (spinnen en krabben) of een 3-delig lichaam (insecten) of ze bestaan uit vele segmenten (Myriapoda (miljoenpoten en duizendpoten)).
Tot de Arthropoda behoren eveneens de beroemde fossiele Trilobieten. Levend tijdens het allereerste begin van de wereldgeschiedenis.
b. Spiralia.
Zo genoemd omdat de eerste delingen van de eicel een karakteristieke (spiraalvormige) klieving vertonen. Daarbij worden 4 macromeren en 4 micromeren gevormd. Daarop komen echter ook uitzonderingen voor!
Tot de Spiralia behoren de
Platwormen (zonder coeloom) en
Rotifera (met ronddraaiende trilharen).
https://upload.wikimedia.org/wikipedia/commons/thumb/6/66/Habrotrocha_rosa_1.jpg/266px-Habrotrocha_rosa_1.jpg
Tot de Spiralia worden gerekend:
a een zeer diverse groep van kleine wormachtige dieren (Gastrotricha etc.).
Zonder coeloom.
b Brachiopoda: nu nog 200 soorten. Veelal levend in diepe zeeën.
Bezitten een lofofoor als krans rond de mond om voedsel te vangen.
Bestaan vanaf precambrium en overheersen tijdens het
Paleozoïcum. Ze bezitten een onder- en bovenschelp i.t.t. de
mollusken die een rechter- en linkerschelp hebben en zich
voeden m.b.v hun kieuwen met daarop trilharen.
c Nemertea: soms zeer lange wormachtige dieren (tot 35 m.)
met een proboscis. Voortbeweging door ciliën. Fossielen
vanaf Cambrium.
d Annelida: de welbekende regenwormen, zeepieren en Nereis.
https://upload.wikimedia.org/wikipedia/commons/thumb/c/cf/Nereis_pelagica.jpg/266px-Nereis_pelagica.jpg
e
Mollusca: met of zonder schelp: de slakken, 2-kleppigen, inktvissen,
chitons.
De Inktvissen komen voor vanaf het Cambrium. Dit geldt eveneens
voor de Anneliden, Mollusken en Slakken.
 Chiton.
Chiton.https://upload.wikimedia.org/wikipedia/commons/thumb/7/70/Cryptoconchus_porosus_%28butterfly_chiton%29.JPG/200px-Cryptoconchus_porosus_%28butterfly_chiton%29.JPG
Conclusie van de schrijvers: de onderlinge relaties .binnen de polyphyletische groep van Spiralia zijn volledig onzeker. Moleculaire studies plaatsen b.v. de Brachiozoa naast de Annelida.
De Mollusca vormen een geheel van onherleidbare groepen. De schelpdieren (2-kleppigen, slakken en inktvissen) zouden een monophyletische groep zijn maar de diverse schelptypen zijn totaal verschillend. De Aplacophora zouden basale mollusken kunnen zijn maar de relatie tussen beide groepen onderling is ingewikkeld.

een
Aplacophora. https://upload.wikimedia.org/wikipedia/commons/thumb/d/d9/Epimenia_verrucosa.jpg/220px-Epimenia_verrucosa.jpg
De Aplacophora hadden geen schelp. Wel naaldvormige spiculae in de huid.
6.1.3 Deuterostomia.
De eerste opening van de blastula ontwikkelt zich tot anus.
Hiertoe behoren de Chordata (waartoe ook de vissen, vogels, zoogdieren en mensen behoren) en de Ambulacralia (waartoe de Echinodermata en de Hemichordata behoren).
a. Hemichordata.
De Hemichordata is een wonderlijke groep waartoe de uitgestorven, oeroude groep van Graptolieten behoort; levend vanaf het Cambrium. Een heel bijzondere groep is de wormvormige Enteropneusta. Sommige celgroepen tonen verwantschap met die bij hogere dieren die de ontwikkeling van hersenen bevorderen.
Ze hebben kieuwaanleg, hart en bloedvaten evenals een dorsale en ventrale zenuwstreng. Al met al worden ze soms beschouwd als voorouders van gewervelden of als overgang tussen gewervelden en ongewervelden. Terloops wordt opgemerkt dat de voorste pharynx bij chordata de aanleg van kieuwen bevat. Bij de hogere vertebraten zijn deze ontwikkeld tot echte kieuwen.
https://upload.wikimedia.org/wikipedia/commons/thumb/f/ff/Enteropneusta.png/220px-
Enteropneusta.png
b. Echinodermata.
Tot de Echinodermata behoren naast zeesterren, zee-egels enz. Ook de tijdens het
precambrium levende Crinoidea (zeelelies)
. Sinds 1864 zijn er enkele nu levende soorten van deze laatste ontdekt op de diepe oceaanbodem.
Echinodermata bezitten een ambulacraal systeem. Volkomen uniek in het dierenrijk!!
Chordata bezitten een dorsale weefselstreng de Notochord; een zenuwstreng gelegen boven de chorda.
De chorda functioneert als endoskelet en ontstaat uit het mesoderm. Ze komt ook voor in de larven van Urochordata.
c. Cephalochordata.
De Cephalochordata worden beschouwd als een belangrijke fossiele groep die nu nog voortbestaat als Amphioxus (lancetvisje). Ze voeden zich door filteren van water dat over de kieuwen stroomt.
Beroemd fossiel: Pikaia.
d. Urochordata == Tunicata.
Alleen in de larven is de chorda en zenuwstreng aanwezig. Ze leefden al tijdens het Cambrium.
Ze zijn een zustergroep van de Craniata. Maar de relatie tussen Cephalochordata, Urochordata en Craniata blijft een groot vraagteken!
6.1.4 Craniata.
Hiertoe behoren de Myxini (slijmprikken en prikken of lampreien). Het zijn Agnatha.
Overigens behoren tot de Craniata alle Vertebrata, inclusief de mens.
a. Gnatostomata.
De kaken bestaan uit beweegbare skeletdelen. Kaken zouden ontstaan zijn uit de 3e en 4e
kieuwbogen.
Dit was nuttig voor een verbeterde waterstroom (?).
De Placodermi waren bepantserde bodemvissen die leefden tijdens Cambrium en vooral het Devoon.
De Chondrichtyes zijn de kraakbeenvissen; haaien, roggen etc.. Ze bestaan al vanaf de vroegste tijden: 430 miljoen jr. geleden. Ze vertonen diverse vormen van voortplanting: ovopaar, ovovivvipaar en vivipaar.
De Actinopterygii zijn de echte, straalvinnige vissen met schubben enz.
De Sarcopterygii worden beschouwd als de stamoudsten van de coelacanthen en de longvissen.
Lobvinnige vissen bestonden al vanaf Devoon, 420 miljoen jr. geleden.
Daarmee schijnt de route naar het ontstaan van dieren met longen en lopend op 4 poten vanzelfsprekend!
b. Amniota.
Nadat de Lissamphibia waren ontstaan via Lepospondyli of Temnospondyli (levend gedurende het Carboon) kwamen hieruit de welbekende kikkers, padden en salamanders voort, zo meent men. Ze komen al voor vanaf het Devoon. Vanwege ontwikkeling van de eieren in het water is de bevruchting normaliter uitwendig en bezitten de eitjes geen schaal.
Als de voortplanting onafhankelijk van water op het land plaats gaat vinden bij reptielen, vogels en zoogdieren is het voorkomen van een amnion vereist die omgeven is door een leerachtige (bij reptielen) of een kalkachtige schaal (bij vogels) of zich zonder schaal ontwikkelt in een baarmoeder.
De Synapsida (alle zoogdieren en hun voorouders) hebben slechts één temporale opening in de schedel.
De Diapsida (de huidige reptielen en vogels) hebben twee openingen in hun schedel.
De Anapsida (zonder temporale schedelopening) zijn uitgestorven. De plaats van de schildpadden is onzeker. Zijn het Anapsiden of zijn hun openingen verdwenen?
De Archosauromorpha omvatten diverse Sauriërs, krokodillen en vogels.
Tot de Protheria of Monotremata behoren de eierleggende zoogdieren: mierenegels en vogelbekdieren.
Ze worden ook wel cloacadieren genoemd. Dus verwant aan reptielen? Maar ze produceren melk!
Alle Theria zijn vivipaar. Hun jongen ontwikkelen zich in het moederlichaam.
Een bijzondere groep naast de Eutheria zijn de Marsupialia (buideldieren). Bij deze dieren verlaat het jong in een zeer vroeg stadium de baarmoeder om zich verder te ontwikkelen in de buidel, samengegroeid met een melktepel. Ze zijn waarschijnlijk ontstaan in de Jura. De eerste fossielen gevonden in het Krijt.
Belangrijk discussiepunt binnen de evolutieleer is warmbloedigheid (homeothermie). Kwam dit al voor bij de Archosauriers en uitgestorven krokodillen? De laatsten hebben een vierdelig hart en een secundair verhemelte.
6.1.5 Holomycota.
Schimmels hebben geen plastiden, wel mitochondriën. Ze vormen mycelium met 2 kernen.
Schimmels ruimen alle organische stoffen op in de natuur samen met bacteriën.
Een tweede zeer essentiële functie van schimmels is de vorming van mycorrhiza rond de wortels van planten; vooral bij bomen en orchideeën. Ze zorgen voor de opname van voedingszouten en benutten organische stoffen die worden uitgescheiden door de wortels. Recent is ontdekt dat de grond onder bossen veel extra CO2 produceert door de activiteit van deze schimmels!
Schimmels leven vaak in symbiose met algen, planten en dieren of ook parasitair.
Op grond van moleculaire gegevens is vastgesteld dat schimmels meer verwant zijn aan dieren dan aan planten. Ook vanwege het feit dat de celwanden bestaan uit chitine, cellulose en glucaan.
Basale groepen zoals de Chitrydiomycota zijn 1-cellig en hebben een zweephaar.
De hogere Fungi hebben secundair gereduceerde flagellen (??).
De Nucleariidae zijn cellen die zich voortbewegen met amoeboïde schijnvoetjes.
De grote subgroep naast de "dieren" vormen de Holomycota (de Fungi).
Want ze zijn 'unikont'.
Chitine verbindt Fungi met de Arthropoda en de basale groepen hebben 1 secundair gereduceerde flagel (unikont).
Maar: de Nucleariidae zijn amoeboïde 1-celligen.
https://upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Nuclearia_sp_Nikko.jpg/220px-
Nuclearia_sp_Nikko.jpg
De Amoebozoa hebben geen vaste celvorm; ze vormen pseudopodia (schijnvoetjes). Amoeboïde cellen komen in diverse supergroepen voor.
Dit alles tezamen genomen maakt het voor mij niet overtuigend dat de Fungi (dus ook de Basidiomycota en de Ascomycota) als nevengroep van de "dieren" dienen te worden beschouwd!
Het aantal flagellen blijkt niet allesbepalend. Amoebozoa (samen met slijmzwammen) vormen wel een heel aparte groep. Met als argument: de flagella zijn gereduceerd.
Alleen de Chytridiomycota vormen cellen met flagel (zoösporen en gameten). De sporen van Microsporidia hebben 2 kernen en een binnenin opgerolde flagel die naar buitenschiet en contact maakt met het epitheel van de darmwand van een gastheer. Kern(en) en cytoplasma worden in de gastheercel geschoten. Een volledig uniek verhaal!!
 Chytridiomycota.
Chytridiomycota.
Ze bezitten geen plastiden. Ze leven wel vaak in symbiose met algen; zoals ook koralen en sponzen.
Ze hebben als gemeenschappelijk kenmerk de synthese van uridine-monofosfaat.
Daarbij zijn 3 genen samengevoegd tot één. Deze zgn. CAD-fusie zorgt voor 1 mRNA voor de synthese van het eiwit.
Ondanks wat de naam zegt kunnen Unikonta 0, 1 of 2 flagellen hebben.
Groepen binnen de Unikonta kunnen heterophyletisch of paraphyletisch zijn.
Misschien stammen Unikonta af van biflagellate vormen.
Hierna volgt een korte bespreking van elke subgroep van Fungi.
a. Microsporidia en Chytridiomycota.
De Microsporidia zijn 1-cellige parasitaire schimmels. Vandaar dat ze geen mitochondriën nodig hebben. Hun algehele bouw is verder aangepast aan de parasitaire leefwijze: geen kinetosoom, centriole, flagella en een sterk gereduceerd genoom.
Daarnaast bezitten ze een zeer specifiek, ingenieus filament dat uitgeschoten kan worden om het darmepitheel te infecteren. Het zijn de kleinste eukaryoten. Met wel een miljoen soorten. Ze zijn ziekteverwekkers bij planten, dieren en mensen. De sporen hebben een soort harpoen waarmee ze organismen binnendringen. Eenmaal binnen vormen ze vaak een plasmodium.
https://upload.wikimedia.org/wikipedia/commons/thumb/3/33/Fibrillanosema_spore.jpg/266px-
Fibrillanosema_spore.jpg
De Chytridiomycota zijn de enige schimmelgroep die voortplantingscellen vormen met een zweephaar.
Anaerobe soorten leven in het spijsverteringskanaal van schapen en vee.
De Chytridiomycota zijn 1-cellig met flagel. Ze zijn meestal saprofyt of parasiet. DNA-onderzoek bewijst dat dit echte schimmels zijn.
https://upload.wikimedia.org/wikipedia/commons/thumb/2/2c/Spizellomycete.jpg/266px-
Spizellomycete.jpg
Sommige soorten vormen draden (hyphae) zonder septa. Die zijn dus meerkernig.
b. Glomeromycota.
De Glomeromycota vormen de mycorrhiza rond plantenwortels. Ze zijn polyfyletisch.
https://upload.wikimedia.org/wikipedia/commons/thumb/c/c7/Gigaspora_margarita.JPG/266px-
Gigaspora_margarita.JPG
Ze vormen clusters of draden die wortels van planten binnendringen. Ze leven in obligate symbiose met mossen, varens en zaadplanten. Ze zijn dus zo oud als het begin van de aardbolgeschiedenis (vanaf 440 miljoen jr. geleden). Deze schimmels zijn niet soortspecifiek en leven samen met 80% van alle landplanten.
Belangrijk: schimmels bestonden al 900 miljoen jr. geleden. Waarmee leefden ze dan samen??
Misschien met cyanobacteriën? Er bestaat een schimmelsoort Geosiphon die in endosymbiose leeft met de cyanobacterie Nostoc. Let wel: endosymbiose!
c. Basidiomycota.
Bouw en voortplanting van deze schimmels zijn in Hoofdstuk 2.3 besproken.
Celwanden met chitine. En daarom in dezelfde supergroep geplaatst als de Arthropoden.
De roesten of Pucciniales (branden) zijn kwaadaardige plantenparasieten. Ze dringen planten binnen via intercellulaire holtes en vormen haustoria. Ook de Ustilaginales zijn obligatoire plantenparasieten. Ze vormen tumorachtige gallen in b.v. Zea (maïs).
d. Ascomycota.
Celwanden met chitine en glucaan.
Truffels, Penicillinum, Pneumocystis (een longparasiet), meeldauwsoorten, Saccharomyces (bakkersgist) en Clasviceps purpurea (het giftige moederkoren in graan) behoren hiertoe.
e. Amoebozoa.
De Amoebozoa vormen een zustergroep van de Ophistokonta, waartoe alle dieren en schimmels behoren.
De Amoebozoa hebben een flagel (meestal secundair gereduceerd) achterwaarts gericht.
Amoebozoa hebben een amoeboïde voortbeweging en geen vaste celvorm. Soms met skelet.
Ze bezitten een actine-myosine-celskelet (vgl. onze spieren).
De ontstaansrelaties binnen de heterogene groepen zijn onbekend.
f. Conosa.
Ook een zustergroep van de Ophistokonta.
Een aantal taxa hebben flagellen.
Hiertoe behoren de slijmzwammen. Ze vormen vruchtlichamen vanuit een meerkernig mycelium.
Ze kunnen ziekteverwekkers zijn.
6.2 Excavata.
De "mond" heeft een specifieke vorm; de zgn. cytosoom. Maar vele groepen missen deze eigenschap of ze is secundair gereduceerd. Een cytosoom komt trouwens ook bij de Ciliata (b.v. pantoffeldiertje) voor.
De flagel is achterwaarts gericht. Groot verschil in aantal flagella: 1, 2, 4 of veel (tot wel 1000).
Sommige groepen zijn amoeboïd qua leefwijze en voortbeweging.
Als ze op anaerobe plaatsen leven zijn de mitochondriën gereduceerd of afwezig. Ze zorgen in het laatste geval nog wel voor de synthese van eiwitten en vetten.
Ze leven grotendeels als commensaal of parasitair.
De Euglenida (groene oogdiertjes) behoren hiertoe. Met of zonder bladgroenkorrels. Met 2 flagellen waarvan er 1 gereduceerd is.
De opslag van reservevoedsel vindt plaats in de vorm van ß 1,3-glucaan.
Dit gebeurt ook in de Haptophyta. Maar die behoren tot de supergroep van Hacrobia.
6.2.1 Metamonada.
Ze hebben 2 kernen en 2 kinetiden (microstructuren voor de beweging van de flagellen). Per kinetide zijn er 4 flagellen; 3 voorwaarts gericht en 1 achterwaarts.
Ze leven allemaal anaeroob en hebben daarom geen mitochondriën. Ze bezitten wel genen voor de aanleg daarvan. Ze leven veelal als commensalen in insecten. Bij de termieten spelen ze een belangrijke rol bij de vertering van lignine en cellulose. Vele soorten zijn pathogeen: b.v. Trichomonas vaginalis die 170 miljoen mensen per jaar infecteert. De infectie behoort tot de zgn. SOA's (seksueel overdraagbare aandoeningen).
6.2.2 Discoba.
Sommige groepen met meer of minder flagellen, andere met amoeboïde vormen van voortbeweging. De laatste soms weer met stadia die wel flagellen hebben.
Bij de Acrasidae vormen zich pseudo-slijmzwammen met vruchtlichamen waarbinnen sporen worden gevormd. Diverse varianten komen bij andere groepen voor. Maar eer zijn steeds kleine verschillen met de slijmzwammen van de Amoebozoa.
a Euglenida.
Voor bouw en structuur zie de pg. 'evolutie-II'.
De Euglenida hebben een pellicula. Dit is een stevige structuur van eiwitten onder de celmembraan die de hele cel spiraalsgewijs omgeeft.
De systematische plaats van verschillende taxa is onduidelijk.
Verondersteld wordt dat de Euglenida afstammen van fagotrofe (heterotrofe) organismen die door endocytobiose aan hun plastiden kwamen.
a 1 Kinetoplastea.
Ze hebben een kinetoplast: een DNA-rijke streng door het hele mitochondrium zo lang als de hele cel. De kinetoplast is verbonden met de basis van de flagel.
Er komen zowel vrij levende groepen voor als parasitaire. De vrij levende met 2 flagellen. De parasitaire soort is o.a. Trypanosoma, de beruchte verwekker van slaapziekte met een ingewikkelde levenscyclus via insecten.
6.3 Archaeplastida.
Zoals eerder opgemerkt: de Archaeplastida vormen een monophyletische groep samen met de cyanobacteriën. Eerste stap(pen): endocytobiose van een blauwalg
Tweede stap(pen): endocytobiose van een eukaryotische alg.
Sommige groepen hebben chlorofyl a, andere chlorofyl a tezamen met phycobillisomen. Deze phycobillisomen bestaan uit een eiwitcomplex die pigmenten bevatten die geel en groen licht kunnen absorberen.
De Viridiplantae bezitten chlorofyl b in de plastiden.
De Streptophyta omvatten zowel een parafyletische groep algen als alle landplanten (Embryophyta).
De landplanten bestaan uit de parafyletische groep mossen en de monofyletische groep van paardenstaarten, varens en zaadplanten.
De Glaucocystophyta vertonen overeenkomst met de cyanobacteriën vanwege peptideglycaan tussen de membranen van de plastiden. Dit is een relict van de celwand van een cyanobacterie (?).
Ze hebben eveneens blaasjes onder de celmembranen. Deze zijn gelijk aan de alveoli van de Alveolata die behoren tot de supergroep van Chromalveolata.
 Glaucocystophyta.
Glaucocystophyta.
Cyanophora paradoxa wordt beschouwd als een levend fossiel vanwege de blijken van primaire endocytobiose als een 'enkelvoudige gebeurtenis' die 1 miljoen jaar geleden plaats greep.
Opm.: je zou deze gebeurtenis toch wel graag eens bewezen willen zien!
Rhodophyta hebben een vergelijkbare celstructuur als de Glaucocystophyta maar met enkele bijzonderheden:
- als enige protist met plastiden bezitten ze fycoerythrine
- het chloroplast-DNA is niet circulair
- reservevoedsel wordt opgeslagen als alpha-1,4-glucaan (vgl. amylopectine)
- er zijn 3 generaties: gametofyt, carposporofyt en tertrasporofyt.
Archaeplastida hebben chlorofyl a, zetmeel als reservevoedsel en cellulose als celwandmateriaal.
De mitochondriën hebben platte cristae.
De Archaeplastida vormen een homophyletische groep binnen de Cyanobateria omdat de eerste stap in hun evolutie bestond in de endocytobiose van een Cyanobacterie.
Daarna kan er endocytobiose hebben plaats gevonden van een eukaryotische alg.
De Rhodophyta hebben vaak generatiewisseling met 3 generaties.
De spermacellen hebben geen flagel maar bewegen zich amoeboïd. Ze worden gevormd in vele gametangia. Terwijl in 1 vrouwelijk gametangium slechts 1 eicel gevormd wordt.
Uit de zygote ontstaat de carposporofyt (2n) die carposporen (2n) vormt. Daaruit groeien tetrasporofyten (2n) die door reductiedeling de sporen (1n) vormen
Hun weefsels bevatten pseudoparenchym en cellulose. Ze hebben zowel chlorofyl a als phycobillisomen en fycoerythrine. Daardoor kunnen ze tot 250 m. diep groeien waar slechts 0.001 % van het daglicht aanwezig is.
Pseudoparenchym komt voor bij roodwieren, schimmels en korstmossen.
Hun cellen bevatten phycobilline voor de fotosynthese. Afkomstig van Cyanobacteriën?
De Glaucocystophyta worden beschouwd als belangrijke overgang vanaf de Cyanobateria omdat ze peptideglycaan bezitten tussen de membranen van de plastiden. De plastiden bevatten chlorofyl a en phycobilline. Dit laatste molecuul maakt opname van licht mogelijk, noodzakelijk voor de fotosynthese. Het bestaat uit 4 pyrrole-ringen. Dit is ook het geval bij chlorofyl maar dan vormen de 4 pyrrolen een ring met een metaal in het centrum.
De Viridiplantae (de planten) vormen een monophyletische groep. Maar ze is zeer divers in morfologie, celstructuur, levenscyclus, voortplanting en biochemie. Veelcelligheid ontstond vele malen onafhankelijk van elkaar! B.v. Volvox. Volvox heeft 2 celtypen: 1 voor de voortplanting en 2 voor de voortbeweging.
De Chlorophyta bezitten chlorofyl a en b, plastiden met 2 membranen, (gevolg van primaire endocytobiose), cellulosewanden en mitochondriën. Met 17.000 soorten komen ze vrijwel overal voor.
Ze zijn zeer veelvormig: met of zonder flagellen, wel of geen kolonievorming, wel of geen celdifferentiatie, vertakt of onvertakt, draadvormig of bladachtige thalli (b.v. zeesla), wel of geen phycoplast-vorming tijdens de celdeling, wel of zonder schaal, met of zonder endosymbiose (met Fungi zoals Lichenen). Soms komt veelkernig weefsel voor, zoals bij Ulvophyceae.
De Streptophyta omvat diverse groepen parafyletische zoetwater groene algen.
De voorlopers (wie waren dat? Wellicht 1-cellige flagellaten met plastiden) van de Streptophyta ontwikkelden de verschillende vormen van landplanten zoals mossen, varens, paardenstaarten en bloemplanten. In de voorbereiding daartoe bezitten ze:
- een celwand van cellulose
- veelcelligheid
- vorming van een phragmoplast tijdens celdeling
- bezit van plasmodesmata
- apicale groei
- driedimensionaal weefsel
- asymmetrische celdeling
- celdifferentiatie.
- biochemische en fysiologische overeenkomsten zoals de vorming van GAPDH (glyceraldehyde-3-
fosfaat-dehydrogenase) onder besturing door het enzym thioredoxine.
Microtubuli van een phycoplast tijdens de celdeling.
Microtubuli van een phragmoplast tijdens de celdeling.
De fylogenetische relatie tussen Streptophyta en landplanten blijft onbekend! De Charophytina (de kranswieren) werden lange tijd beschouwd als voorlopers van de landplanten maar dit blijkt op grond van moleculaire gegevens niet te kloppen!
Embryophyta met
1. Bryophyta (mossen).
De Bryophyta zijn hierboven al uitvoerig besproken. Ze leefden vanaf Devoon/Carboon.
2. Rhyniophytina (b.v. Cooksonia) leefden vanaf Siluur/Devoon. De stelen zijn bladerloos en dichotoom vertakt. Ze zijn isospoor. De sporophytgeneratie overheerst. Ze kwamen wereldwijd voor. De eveneens uitgestorven nauwverwante groep Zosterophyllopsida (b.v. Asteroxylon) vormen een parafyletische groep. Eveneens dichotomie en isosporie. Ze hebben exarch xyleem.
3. Lycopodiopsida (wolfsklauwen) zijn isospoor en de sporofyllen zijn dichotoom vertakt. Afwisselend microphylli met en zonder sporangium. Ondergrondse prothallia leven in symbiose met Fungi.
Ze bezitten floëem.
4. Selaginellales (b.v. Engels mos) komen nu nog overal voor met ongeveer 700 soorten. Bij de Selaginellales vinden we megasporangia en microsporangia. De mannelijke gametofyt blijft in de spore en de vrouwelijke gametofyt groeit uit de spore.
5. Lepidodendrales groeiden uit tot hoge bomen tijdens het Carboon. O.a. Sigillaria.
Sporangia groeien op een sporofyl die samen een strobilus vormen.
6. Isoetales (biesvarens) zijn veelal aquatisch. Met megasporofyllen en microsporofyllen. Isoetales bezitten cambium.
7. Monilophyta (varens). Met 300 geslachten en 9000 soorten. Wereldwijd verspreid en vooral in schaduwrijke plaatsen. Vaak met wortelstokken Varens hebben primaire plastiden.
Het studieboek van Jens Boeningk c.s. neemt de Monilophyta samen met de Spermatophyta (zaadplanten) als twee zustergroepen: de Euphillophyta (letterlijk de bladplanten). De Monilophyta bevatten dan de volgende groepen:
a Marattiopsida, het sporangiumwand bestaat uit 2 cellagen (eusporangiaat).
Dit is ook het geval bij b en c. Ze bestaan vanaf Carboon. Antheridia en
archegonia verzonken in de onderzijde van het prothallium.
b Psilopsida, o.a. Ophioglossales (b.v. Addertong met 2 typen bladeren;
één met alleen sporangia. Psilotum zonder wortels, maar
wel wortelstokken en dichotoom vertakte stengels (vgl. Rhynia)..
Het prothallium leeft ondergronds, samen met Fungi.
c Equisetopsida ( de paardenstaarten). Tot 30 m. hoge boomfossielen
(Calamitaceae) in het Carboon.
d Polypodiopsida (echte varens). De sporangiumwand bestaat uit
1 cellaag (leptosporangiaat).
Boomvarens tot 20 m. hoog in het Carboon. Hiertoe behoren ook
de Salviniaceae (de vlotvarens, waaronder Azolla, watervarens).
Ze zijn heterospoor (met macro- en microsporen) en dioece (ze
hebben mannelijke en vrouwelijke prothallia)
De varens vertonen concentrische bouw van de vaatbundels met xyleen binnenin en phloeem daar omheen.
Vergelijk met 1- en 2-zaadlobbige zaadplanten:
Een dwarsdoorsnede van een 1-zaadlobbige plant
Een dwarsdoorsnede van een varenbladsteel.
Voor de verdere bouw van varens en paardenstaarten zie Hoofdstuk 2.5.2 en Hoofdstuk 2.5.3.
Gymnospermen.
De voorlopers van de Gymnospermen zouden de uitgestorven Progymnospermen zijn. Zij vormden geen zaden maar bezaten wel cambium dat secundair xyleem en secundair floëem vormde.
Gymnospermen waren in eerste instantie houtachtige planten. Kruidachtige Gymnospermen zouden daaruit later zijn ontstaan.
Bij de Gymnospermen is de gametophyt sterk gereduceerd. Dit is besproken in Hoofdstuk 2.6.1
Tot de Gymnospermen worden gerekend:
1. Cycadopsida: de Palmvarens die zeer uitgebreid voorkwamen in het
Mesozoïcum. Nu vooral in tropische gebieden met 300 soorten. Ontstaan
tijdens Carboon. Spermatozoïden met flagel.
2. Ginkgoöpsida: Ook ontstaan in Carboon en wijd verspreid in
Mesozoïcum. Dichotome bladnervatuur. Nog steeds beschouwd als
oorspronkelijk? Levend fossiel? Spermatozoïden met flagel.
3. Cordaitopsida: Uitgestorven. Op noordelijk halfrond verspreid
tijdens Paleozoïcum.
4. Coniferopsida: levend vanaf Carboon. Met naaldvormige of
schubvormige bladeren. Hiertoe behoren de Gnetales: de
systematische plaats hiervan is erg onduidelijk. Nauw verwant aan Pinus?
Of aan "bloemplanten"? De Gnetales omvatten drie nu levende
plantengroepen: Gnetum, Welwitschia en Ephedra.
5. Glossopteridopsida: bomen die leefden tijdens Perm/Trias.
6. Benettitopsida: leefden gedurende Trias tot Krijt.
Met "bloemachtige" kegelstructuur.
Magnoliopsida.
De Magnoliopsida zijn Angiospermen (bedektzadigen).
Kenmerken van de Angiospermen zijn:
- bezit van zeefvaten (floëem) met begeleidende cellen
- meeldraden met 2 helmhokken
- de mannelijke gametofyt bestaat uit 3 cellen
- er vindt dubbele bevruchting plaats en daardoor vorming van triploïd endosperm (reservevoedsel)
De Magnoliopsida bestaan vanaf vroeg-Jura.
Met 230.000 soorten vormt ze de meest diverse groep van landplanten!
Tot de Angiospermen behoren de Monocotylen en de Dicotylen tezamen met een aantal basale groepen: de ANITA-clade.
De basale groepen en de Monocotylen hebben stuifmeelkorrels met slechts 1 opening. De Eudicotylen hebben stuifmeelkorrels met 3 openingen.
De systematische plaats en de fylogenie van de Magnoliopsida is onduidelijk.
Over de verdere indeling van de Magnoliopsida bestaat veel verschil van inzicht.
Tot de Magnoliopsida behoren:
1. Basale Angiospermen: bloemdelen afwisselend of drievoudig
gespiraliseerd, meestal geen vruchten.
Het zijn vooral houtachtige planten. Met 8600 soorten.
- ze vormen geen monophyletische groep.
- de idioblasten bevatten etherische oliën.
- de voortplantingsorganen zijn meestal gerangschikt in een spiraal.
- de vruchtbeginsels zijn meestal niet vergroeid.
- ze bezitten primaire plastiden met chlorofyl a en b
- het aantal kernen in de embryozak varieert van 4-8-9.
O.a. de Nymphaeales (o.a. de waterlelie) worden hiertoe gerekend. Ze bezitten 4 kelkbladeren (sepalen) en wel tot 50 kroonbladeren (petalen). Gerangschikt in spiralen.
2. Magnoliida: meeldraden staan in spiralen. O.a. Piperales, Laurales en Magnoliales
3. Monocotylen: o.a. lelies, grassen, orchideeën en bolgewassen.
- de zaden hebben 1 cotyl (zaadlob)
- ze hebben bijwortels
- de vaatbundels zijn collateraal gesloten
- de vaatbundels liggen verspreid (zie boven)
- de bladeren staan verspreid
- de bladeren zijn rolrond of paralelnervig.
4. Ceratophyllales: de hoornbloemfam. Op vele manieren ondergebracht
in een systeem; instaande tussen Monocotylen en Chloranthales?
Stuifmeel zonder opening. Geen trachaea-xyleem.
V.b. van convergente evolutie?
5. Eudicotylen:
- de zaden hebben 2 cotylen
- stuifmeelkorrels met 3 openingen
- ze hebben hoofd- en zijwortels
- de vaatbundels collateraal-open
- de bladeren staan verspreid, tegenover elkaar, kransvormig of alternerend
- bladnervatuur: hoofdnerf met zijnerven.
Hiertoe behoren:
de Rosiden: vrije kroonbladeren en 2 rijen meeldraden;
embryozak met 2 integumenten
de Asteriden: grote bloemkroonbladeren, minder meeldraden dan
bloemkroonbladeren, embryozak met 1 integument.
Basale Eudicotylen met Ranunculales, Sabiales, Proteales, Trochodendrales, Buzales.
o.a. de familie Ranunculaceae. Bloemen radiair symmetrisch; soms zygomorf.
Hiermee hebben we het hele rijk van de Angiospermen gehad. De indeling in vele orden en families is deels gebaseerd op moleculaire gegevens en morfologische kenmerken. De systematische plaats van orden en vooral van families vertoont vele verschillen in criteria en opvattingen. Er is vrijwel niets te zeggen over periode van ontstaan van de verschillende orden en families.
Kenmerken als symmetrie, wel of niet vergroeidbladig, onderstandig of bovenstandig vruchtbegingel, bladstand, nervatuur, bloemstructuren etc. variëren veelal binnen elke orde.
Indelingsbegrippen als rijken, (hoofd)afdelingen, klassen, orden, families etc.) zijn hiermee als achterhaald beschouwd.
6.4 Rhizaria.
Rhizaria is een erg diverse supergroep en zeer algemeen. De meeste taxa hebben diverse typen pseudopodia.
De Retaria hebben een skelet van kalk of kiezel (Foraminiferen en Radiolariën).
Ze bewogen zich voort m.b.v. pseudopodia. Ze komen voor vanaf het begin van de aardgeschiedenis.
De Chlorarachniophyta hebben chloroplasten! Deze hebben ze waarschijnlijk verkregen door endocytobiose van een groenwier of een roodwier. Bewijs: tussen de membranen van de plastide bevindt zich een nucleomorf. Dit zou een overblijfsel zijn van de kern van het groenwier. De zoösporen kunnen een flagel hebben!
Bij de Cercozoa komen zowel exemplaren voor met schijnvoetjes als met flagellen.
Binnen deze groep heeft Paulinella een chromatofoor die afkomstig is van een cyanobacterie.
https://upload.wikimedia.org/wikipedia/commons/thumb/c/ca
/Paulinella_chromatophora.jpg/220px-Paulinella_chromatophora.jpg
Als je alles wilt weten over de bewijsvoering rondom deze veronderstellingen lees dan op de volgende site:
6.5 Chromalveolata (Alveolata en Stramenopiles).
De Alveolata hebben alveoli onder de membraan. Ze hebben flagellen (b.v. de Dinophyta) of trilharen (b.v. Paramecium). Ze bezitten wel of geen bladgroenkorrels.
De relaties binnen de groepen als het gaat over de herkomst van plastiden is zeer ingewikkeld. Moleculaire studies vertellen de gemeenschappelijke afkomst van alle plastiden die afkomstig zijn van Rhodophyta (roodwieren). Toegegeven wordt dat endocytobiose een zeer complex proces is dat de werking van vele genen en de route van vele ingewikkelde stofwisselingsprocessen betreft.
Had de voorouder van Alveolata, de Stramenopiles en de Rhizaria (de SAR-clade) nu wel of geen plastiden. Kreeg de voorouder een plastide door secundaire endocytobiose en verloren vele afstammelingen deze weer? Of ontstond secundaire endocytobiose vele malen? Of verkreeg een subgroep de plastide van een roodwier dankzij secundaire endocytobiose en gaf deze de plastide door aan andere cellijnen via tertiaire endocytobiose?
Vast staat: alle plastiden van alle groepen zijn aan elkaar verwant.
En: de Dinophyta bestaan vanaf het Cambrium.
Je ziet: alles is mogelijk en weinig is zeker!
6.5.1 Ciliophora
Hiertoe behoort het ons welbekende pantoffeldiertje. Met haar macro- en micronucleus. Bovendien duizenden ciliën die in een schitterend ritme de doelgerichte voortbeweging en voeding mogelijk maken. Ze bezitten geen plastiden!
6.5.2 Dinophyta.
Deze groep van eukaryotische 1-celligen vertoont de meest voorkomende eigenschappen van alle eukaryota. Sommige van de Dinophyta produceren licht. Evenals vuurvliegjes en vele diepzeevissen. Lichtproductie is mogelijk doordat een eiwit luciferine door een enzym luciferase aangezet wordt om licht te geven.
1 plastide 2 thylakoid 3 mitochondrium 4 Golgi apparaat 5 pyrenoid
6 kern
7 sulcus 8 celluloseplaten.
Dinophyten bezitten secundaire plastiden met 3 membranen + chlorofyl a en c.
Of ze hebben tertiaire plastiden met 4 membranen + chlorofyl a en c of chlorofyl a en b.
Ze kunnen zowel autotroof als heterotroof zijn. Ze bevatten ook mitochondriën.
Ze bezitten 2 flagellen: 1 loopt via een longitudinale groeve naar buiten,
1 loopt door een equatoriale groeve. Deze heeft fijne ciliën.
De cellen bezitten wel of geen cellulose-platen.
Er vindt zowel geslachtelijke als ongeslachtelijke voortplanting plaats.
Heel bijzonder: het DNA in de kern bevat geen histonen.
Dinophyta bestaan al sinds het Cambrium; dus al vanaf het begin van de wereldgeschiedenis!
Er zijn 4000 (!!) soorten.
Ze kunnen in symbiose leven met Radiolaria, Foraminifera, Mollusken, Cnidaria en Ciliata.
Sommige zijn parasitair.
Dinophyta kunnen over honderden vierkante km's algenbloei veroorzaken. Ze produceren daarbij giftige stoffen waaraan andere waterorganismen bezwijken.
De originele plastiden zijn van het Peridinium-type. Ze zijn afkomstig van Rhodophyta (roodwieren). Deze plastiden hebben 3 membranen, thylakoiden in paren van 3 en chlorofyl a + c. Ze bezitten verder ß-caroteen en xantophyl. ß-caroteen komt voor in groente, fruit en micro-algen. Het is een provitamine voor vit. A.
https://upload.wikimedia.org/wikipedia/commons/thumb/7/7c/Granumstructuur.png/432px-Granumstructuur.png
Xanthine is een purine-derivaat en kan worden omgezet in guanine; een belangrijk element van de nucleotiden (kernzuren) die noodzakelijk zijn voor de bouw van DNA en RNA.
6.5.3 Apicomplexa.
De Apicomplexa zijn zeer bijzonder gebouwde 1-celligen en zijn obligate parasieten. Een berucht v.b. is de malariaparasiet. Ze hebben 1, 2 of 3 generatiewisselingen via verschillende gastheren. Met diverse vormen van geslachtelijke voortplanting. Ze maken gebruik van een apicoplast, afkomstig van een chloroplast die omgeven is door 4 membranen. Dus binnen gekomen door secundaire endocytobiose. Ze speelt nog wel een rol in de biosynthese van vetzuren en isoprenen.
Plasmodium, de verwekker van malaria behoort tot de
Apicomplexa. Apicoplexa
Apicoplexa
1. polaire ringen 2. microtubuli 3. micronemen (deel van Golgi netwerk) 4. klierorganellen 5. alveoli-complex 6. apicoplast met 4 membranen 7. kern 8. mitochondrium 9. Golgi netwerk 10. microtubuli.
 Chrysophyceae.
Chrysophyceae.1. flagel met tripartite haren 2. korte flagel 3. Golgi netwerk 4. mitochondrium 5. kern 6. plastide
7. thylakoid 8. endoplasmatisch reticulum.
6.5.4 Stramenopiles.
De Stramenopiles vormen een zeer gevarieerde zustergroep van de Alveolata.
Ze is gevarieerder dan alle Metazoa (dieren) en Embryophyta (planten) samen!
Ze hebben gemeenschappelijk: een voorste flagel met holle, driedelige Mastigonema (haren op de flagellen).
Vele groepen hebben chloroplasten; vele andere niet. Of sterk gereduceerd chlorofyl. De groepen zelf zijn wel monofyletisch maar hun onderlinge relaties zijn volstrekt onduidelijk.
De groepen binnen de plastidenloze Stramenopiles verschillen enorm van elkaar: 1 flagel, 2 flagellen of vele. Ze zijn 1-cellig of kolonievormend.
De groepen met chlorofyl worden beschouwd als monophyletisch.
Wel zijn daarbinnen drie hoofdgroepen met verschillende manieren voor de vorming van carotenoïde.
6.5.5 Perenosporomycetes (Oomycetes).
Ze leven veelal parasitair. 400 soorten. Ze zijn veelal 1-cellig en sommige zijn veroorzakers van ernstige plantenziekten; b.v Phytophtora infestans (aardappelziekte).
De interne classificatie blijft vele vragen oproepen.
6.5.6 Phaeophyceae.
Bruinwieren. 1800 soorten. Tot 60 m. lange kelp; levend in zee.
De voortplantingscellen met 4 flagellen: 1 voor, 1 achter, 2 lateraal. Een van de twee met 2 rijen tripartite, driedelige haren.
Eén plastide per cel met 4 membranen en chlorofyl a, c1 en c2. Plus de hulppigmenten ß-caroteen, fucoxanthine, diadinoxanthine en diatoxanthine.
Met als reservevoedsel Chrysolaminarine (een polysacharide).
Een haploïde en een diploïde generatie.
Ze hebben verschillende soorten weefsels. Vaak met luchtblazen De celwanden bestaan uit cellulose samen met zouten van ingewikkelde verbindingen die samen algininezuur (een mengsel van polyuronzuren) vormen. De Phaeophyceae (de bruinwieren), de Chrysophyceae (de gouden wieren) en de Bacillariophyceae (de al eerder besproken diatomeeën) vormen samen de Ochrophyta. Samen met o.a. de Oomycetes vormen ze de Stramenopiles.
6.5.7 Chrysophyceae.
Goudalgen. Met of zonder chlorofyl. Of sterk gereduceerde plastiden. Eén of
veelcellig. 2 flagellen. Dankzij moleculaire studies samengebracht in 1 groep.
6.5.8 Bacillariophyceae.
Diatomeeën.
De cellen bezitten chlorofyl a en c + fucoxanthine. Het zijn secundaire plastiden met 4 membranen.
Reservevoedsel wordt opgeslagen in de vorm van chrysolaminarine. Een polysacharide die is
opgelost in een vacuole. Soms is olie het reservevoedsel; dit is ook nuttig voor drijfvermogen.
Diatomeeën leven in een siliciumdoosje-dekseltje. Er is geslachtelijke voortplanting en deze is vereist om terug te keren naar de oorspronkelijke grootte. De cellen met silicaatwanden zijn bedekt met
een organische laag.
Sommige gameten hebben een flagel met tripartite haren.
Diatomeeën zijn de enige algencellen met een dubbel stel chromosomen (2n) in al hun cellen;
uitgezonderd de gameten.
Diatomeeën verschenen, zoals al eerder vermeld, pas in de Jura. Veel te laat dus. Diatomeeenaarde wordt nuttig toegepast in insecticiden, tandpasta en wegmarkeringen.
De schrijvers geven wel toe dat de moleculaire verwantschapsgeschiedenis en de diverse soorten plastiden bij deze organismen moeilijk (of niet) tot elkaar te herleiden zijn.
Tertiaire endocytobiose wordt zo nodig te hulp geroepen, evenals zeer vaak herhaalde secundaire endocytobiose.
We krijgen hierbij te maken met een essentieel uitgangspunt voor de hele indeling van alle organismen. Er zou sprake geweest zijn van endocytobiose tijdens het ontstaan van planten en dieren.
Endocytobiose van cyanobacteriën om het proces van anaerobe fotosynthese mogelijk te maken,
endocytobiose van diverse soorten bacteriën om mitochondriën in alle mogelijke weefsels te verklaren,
endocytobiose van diverse groene algen of roodwieren om te zorgen voor fotosynthese in de vele typen groene planten.
De theorie begint met de aanname dat de celkern zou kunnen zijn ontstaan doordat een ringvormig DNA, zoals voorkomt bij bacteriën, zich vasthechtte aan de celmembraan of celwand.
Vraag: hoe kwam dit DNA naar binnen? Door fagocytose? Maar dan wordt het volledig afgebroken.
Kernmembraan en endoplasmatisch reticulum zijn, zo meent men, ontstaan door instulping.
Vraag: hoe werden de duizenden (miljoenen) microsomen daar zo netjes op "gemonteerd"?
Aerobe bacteriën als Rickettsia evolueerden tot mitochondriën.
Vraag: is dit door experimenten in laboratoria bevestigd?
Cyanobacteriën (foto-autotroof) ontwikkelden zich tot chloroplasten.
Vraag: is deze mogelijkheid onderzocht in laboratoria?
Secundaire endocytobiose kwam voor als een heterotrofe eukaryoot een groene algencel opnam. De aanwezigheid van een nucleomorf in een chloroplast zou hiervoor bewijs zijn.
Opm.: dit zou aanleiding kunnen zijn tot diepgravende experimenten.
https://th.bing.com/th/id/OIP.Tb99dCyBAMgb6TUpgYmOcgHaDy?w=349&h=178&c=7&o=5&pid=1.7
Een nucleomorf wordt beschouwd als een restant van de kern van b.v. een alg die door secundaire endocytobiose is opgenomen in een andere cel.
Deze theorieën zijn noodzakelijk om het ontstaan van eukaryote dieren en planten te begrijpen.
Bladgroenkorrels en mitochondriën zijn zo ingewikkeld gebouwd en zo specifiek georganiseerd om hun essentieel-noodzakelijk functies te vervullen, dat elke theorie omtrent hun ontstaan tekort schiet!
Het zal duidelijk zijn dat door uit te gaan van endocytobiose het ontstaansprobleem slechts wordt verschoven. Bovendien gaat deze theorie voorbij aan het algemeen bekende feit dat celvreemde eiwitten nooit zullen worden geaccepteerd door een organisme. Een lichaamsvreemd eiwit wordt onmiddellijk herkend en afgebroken door enzymen.
Symbiose komt zeer veel voor, endocytobiose is een (voorzover mij bekend) aan de natuur vreemde vooronderstelling! De begrippen endosymbiose en endocytobiose worden vaak met elkaar verward!
6.6 Hacrobia.
Haptophyta vormt wellicht een zustergroep van de SAR-clade (Alveolata, Stramenopiles en Rhizaria).
De Cryptophyta zijn misschien meer verwant aan de Archaeoplastida.
Dus de fylogenetische positie is geheel onduidelijk. Daarom wel genoemd: 'incertae sedis eukaryoten'.
Sommige soorten hebben chlorofylkorrels, andere niet.
Sommige hebben 2 flagellen, soms als haptonema om zich vast te hechten. De subgroep Tolonema heeft een ingewikkelde skeletstructuur die bestaat uit microtubuli en microfilamenten. Een andere subgroep heeft alveoli onder de celmembraan en tripartite haren op de lange flagel (vgl. de Chrysophyceae).
6.6.1 Haptophyta.
De Haptophyta, met 500 soorten, kunnen enorme velden met algenbloei veroorzaken met vissterfte als gevolg. Op aarde zijn ze bijna alomtegenwoordig en vormen een groot deel van alle biomassa.
De cellen van de
Haptophyta zijn bedekt met celluloseplaten. Deze zijn soms versterkt door calcium. Daardoor vele fossielen in
Carboon en
Krijt.Ze bezitten secundaire plastiden met 4 membranen en bevatten chlorofyl a en c + fucoxanthine en diatoxanthine. Ook hier is het endoplasmatisch reticulum een voortzetting van de chlorofylmembranen net als bij de Chrysophyceae.
Er is een haptonema tussen beide flagellen. De haptonema heeft een zeer bijzondere structuur met 6 of 7 microtubuli.
Reservevoedsel wordt eveneens opgeslagen als chrysolaminarine. Soms ook als olie of paramylon (net als bij de Euglenida). Paramylon is een ß-1,3 polymeer van glucose.
Sommige soorten hebben een oogvlek (vgl. Euglena).
6.6.2 Cryptophyta.
De Cryptophyta, met 200 soorten. Deze laatste leven bij lagere lichtintensiteiten en dus in dieper water.
Ze hebben soms wel, soms geen bladgroenkorrels. Bezit van flagellen en de structuur van celwanden kunnen onderling verschillen. De onderlinge relaties zijn onduidelijk.
Chlorofyl a en c2 + fucoxanthine en diatoxanthine afkomstig van endocytobiose van een roodwier. Zetmeelkorrels en een nucleomorf (beschouwd als restant van de roodwierkern) bevinden zich tussen de 2e en 3e membraan. Zelfde structuur van het endoplasmatisch reticulum als bij de Haptophyta.
Andere groepen 1-celligen leven als endosymbionten samen met Cryptophyta.
Cryptophyta hebben geen celwand. In plaats daarvan een drielagige periplast die bestaat uit 2 lagen eiwitplaten met daartussen een derde eiwitmembraan.
Een lange flagel met 2 rijen stevige bipartite haren; de kortere flagel heeft slechts 1 rij haren. Een ander geslacht heeft slechts haren op 1 van de 2 flagellen.
Vele fossielen van de Cryptophyta bezaten geen plastiden.
De belangrijkste groeve van de cellen is van binnen bezet met explosieve organellen. Daarnaast zijn injectosomen met vergelijkbare structuur verspreid aanwezig onder de periplast. Bij prikkeling stoten ze een dubbele opgerolde draad naar buiten. Waarschijnlijk als verdediging tegen vijanden.
In de zgn. SAR-clade worden de Hacrobia samengenomen met de Chromoalveolata en de Rhizaria. omdat ze chlorofyl c bezitten.
De systematische plaats van deze groepen is echter geheel onduidelijk. De Haptophyten zouden een zustergroep van de SAR-clade kunnen zijn of meer verwant aan de Archaeoplastida (waartoe ook de Chlorophyten en de Embryophyten behoren).
Hoofdstuk 7 Megasystematiek.
Nu we tot zover gekomen zijn is het goed om te zien wat de uitgangspunten zijn om te komen tot het hier boven besproken cladistische systeem. Vooral ook omdat de indeling van de eukaryoten de afgelopen 30 jaar enorm is veranderd. Moleculaire gegevens sinds 1980/1990 hebben de oorspronkelijke morfologische criteria voor indeling in sterke mate verdrongen. Vooral de gedetailleerde kennis van min of meer complete genomen hebben bijgedragen aan de fylogenetische kennis van relaties tussen groepen organismen. De basenvolgorde in DNA en RNA is daarbij het belangrijkste criterium.
Het dient daarbij te gaan om meetbare fylogenetische relaties om de evolutie van organismen tijdens de aardgeschiedenis op te sporen.
De evolutiegedachte zelf is bijna zo oud als de mensengeschiedenis.
In India bestonden overoude ideeën rond het ontstaan van alle levende wezens uit oeroude voorouders.
De Griek Anaximander leefde ongeveer 600 v. Chr. en dacht na over het ontstaan van de mens uit andere levende wezens. Bovendien ging hij uit van 'spontane generatie'. Dit laatste "geloof" heeft de geschiedenis van de "wetenschap" beheerst totdat de natuurgeleerde Pasteur (1822-1895) aantoonde dat dit niet mogelijk is.
Jean Baptiste Lamarck (1744-1820) meende te kunnen bewijzen dat verandering van leefomgeving of hevige concurrentie om ruimte en voeding veranderingen in de bouw van organismen kon teweeg brengen dankzij herhaalde oefening. De nieuw verkregen eigenschappen zouden erfelijk kunnen worden.
Charles Darwin en Alfred Russel Wallace bouwden hierop voort en beschreven de 'natural selection' als de drijvende kracht achter het ontstaan van nieuwe soorten. Het boek van Darwin,
The origin of species ....... (1859) hoop ik later wat uitvoeriger te bespreken..
Darwin greep sterk terug op een boek van Lyell: Principles of geology.
Dit boek ging uit van zeer geleidelijke geologische processen, vergelijkbaar met huidige veranderingen door erosie, sedimentatie en ontstaan van hoogteverschillen. Aardbevingen en vulkaanuitbarstingen speelden alleen zijdelings een rol.
7.1 Fylogenetische bomen.
Fylogenetische bomen gaan uit van gemeenschappelijke kenmerken binnen groepen die onafhankelijk van elkaar ontstaan zijn (convergentie) of terug gaan op een enkelvoudige oorsprong (homologie).
Beide zijn vaak moeilijk van elkaar te onderscheiden. Goede v.b. van convergentie zijn succulentie binnen geheel verschillende plantenfamilies en bezit van zwempoten bij diverse families van watervogels.
Synapomorfieen zijn nieuw ontwikkelde eigenschappen door een voorouder.
De volgende twee begrippen kunnen niet dienen als hulpmiddelen bij het opstellen van fylogenetische lijnen:
- Symplesiomorfieën gaan terug op vroeger verkregen eigenschappen.
- Autopomorfieën vinden slechts plaats in 1 taxon.
De zustergroepen binnen claden kunnen gekarakteriseerd worden door specifieke synapomorfieen.
Een monofyletische groep kan worden teruggevoerd op 1 voorouder.
Een parafyletische groep gaat terug op dezelfde voorouder als de doelgroep maar behoort niet tot dezelfde doelgroep.
Polyfyletische groepen hebben dezelfde of sterk vergelijkbare eigenschappen maar hebben geen gemeenschappelijke voorouder.
Je kunt verschillende soorten berekeningen hanteren om de takken van een cladistische boom te "berekenen". Bekend zijn de 'bootstrap method' en de 'jack-knifing method'.
Alleen de eerste geeft redelijk betrouwbare (?) uitkomsten. Daarbij worden willekeurig gekozen data uit een groep organismen gecombineerd met nieuwe (?) data. Daaruit wordt een nieuwe 'tak' berekend (?). Vervolgens wordt berekend hoe vaak de monofyletische groep voorkomt in de nieuwe 'takken'.
De frequentie waarbij de nieuwe clade als monofyletische groep wordt beschouwd is de 'bootstrapwaarde' in % (?).
Wie dit precies begrijpt mag het zeggen!
Ik vermoed dat bij deze "berekeningen" allerlei vormen van willekeur of 'doeldenken' op de loer liggen.
Dit geldt wellicht nog meer voor de hierna te bespreken methoden voor de bepaling van fylogenetische evolutielijnen.
Aan de basis daarvan ligt de techniek van Polymerase Chain Reaction (PCR). Met deze techniek worden vele kopieën gemaakt van een stukje DNA.
Daarna worden homologe nucleotiden of proteïnen in een matrix geplaatst. Op grond daarvan worden paren van taxa genomen en hun onderlinge afstand (?) bepaald. Verschillende aldus verkregen taxa worden geïntegreerd en omgezet in een patroon van 'takken'.
Het kan ook op een andere manier: neem de gemiddelde afstand (?) tussen taxa en voeg daarbij de expressie (?) van elk kenmerk samen met het veronderstelde minimale aantal (?) mutaties dat nodig is voor de berekening (?) van de 'boom'.
Dankzij deze methodes kan een cladogram (een patroon van 'takken') ontstaan of een fylogram (gemeten moleculair bepaalde afstand tussen 'takken' = de fylogenetische verandering in de tijd).
Wie dit allemaal kan volgen; graag een reactie!
Er kan nog gebruik gemaakt worden van UPGMA (Unweighted Pair Group Method with Arithmetic mean) of Neighbour -joining methods.
UPGMA gaat er vanuit dat de evolutiesnelheid constant is. Opm.: Een merkwaardige veronderstelling!
Neighbour-joining methods zijn gebaseerd op 'minimale evolutie en houden rekening met verschil in snelheid (?) van evolutie.
Daarnaast zijn er nog methoden die uitgaan van maximale zeldzaamheid of maximale waarschijnlijkheid van voorkomende veranderingen. De eerste gaat er vanuit dat evolutie de kortste (?) weg neemt. De tweede kiest voor de 'boom' die de hoogste waarschijnlijkheid heeft op grond van het waarschijnlijkste (?) evolutiemodel.
Nu blijkt de moleculaire diversiteit binnen de Metazoa (de "dieren") en binnen de Archaeplastida (de "planten", de groenwieren en de roodwieren) veel lager te zijn dan die bij Amoebozoa, Rhizaria, Excavata en Chromalveolata.
De diversiteit kan worden gemeten door het tellen van het aantal nucleotiden. Daarbij moet rekening worden gehouden met het aantal gesubstitueerde nucleotiden. Substituties binnen triplets die coderen voor aminozuren zijn minder waarschijnlijk. Vele modellen werden hiervoor ontwikkeld en deze werden steeds complexer dankzij de ontwikkeling van steeds machtiger computers.
Het General Time Reversible (GTR) model gaat uit van de relatieve waarschijnlijkheid (?) van verandering van elke nucleotide.
Het Junkes-Cantor (JC) model is het eenvoudigst. Het gaat er vanuit dat elke substitutie even waarschijnlijk is en dat de 4 basen (A, T, C,en G) evenveel voorkomen. Voor beide aannames geldt
m.i. een grote mate van onwaarschijnlijkheid.
7.2 Eigenschappen van de drie domeinen:
Bacteria: Archaea:
prokaryoot prokaryoot
circulair chromosoom circulair chromosoom
70S-ribosomen 70S-ribosomen
- introns tussen de genen
anders RNA-polymerase en transcriptiefactoren
vergelijkbaar
membraanlipiden vetzuur-esters membraanlipiden isoprenoïden
Eukaryota:
celorganellen met membranen
kern met lineaire chromosomen
70S of/en 80S-ribosomen
introns tussen de genen
RNA-polymerase en transcriptiefactoren vergelijkbaar
membraanlipiden vetzuur-esters.
Opm.: isoprenen zijn 2-methyl-1,3-butadiënen.
Het is belangrijk hierbij te bedenken dat genen en chromosomen zich kunnen verplaatsen tussen de drie domeinen.
Een grote vraag blijft hoe de drie domeinen evolutionair gezien met elkaar samenhangen.
De vaak extreme omstandigheden waaronder de Archaea leven zouden oorzaak geweest kunnen zijn van snelle evolutie.
Een gemeenschappelijke voorouder van de drie domeinen bestaat niet; ook niet in theorie!
Bacteria.
Er zijn 4 groepen bacteria die in staat zijn tot anoxygene fotosynthese. Dat wil zegen dat ze in staat zijn tot fotosynthese maar zonder O2 te produceren.
Alleen de cyanobacteria zijn in staat tot oxygene fotosynthese. Zij produceren dus zuurstof bij dit proces. Zij moeten daarom wel de oorsprong zijn van alle plastiden in het "plantenrijk".
De 5 groepen Proteobacteria nemen een bijzondere plaats in. Want het geslacht Rickettsia bevat leden die het dichtst staan bij mitochondria. Vandaar de theorie dat alle mitochondria dankzij endocytobiose zijn ontstaan vanuit deze groep bacteria.
Archaea.
Soms mist de celwand helemaal en is dan vervangen door laagje glycoproteïnen.
tRNA's hebben vaak gemodificeerde basen, vooral
archaeosine i.p.v.
guanine.
vgl.
 archaeosine guanine
archaeosine guanine Het archaeosine is waarschijnlijk van belang om hoge temperaturen te kunnen doorstaan.
Er zijn een vijftal methaan producerende groepen.
Eukarya.
Primaire plastiden hebben 2 membranen en bezitten chlorofyl a of a + b.
We vinden ze in Archaeplastida (de Glaucosystophyta, de Rhodophyta, de Chlorophyta en de Streptophyta). De Streptophyta omvatten alle landplanten. De plastiden zijn afkomstig van cyanobacteria via het proces van endocytobiose.
Secundaire plastiden hebben 3 of 4 membranen. De Alveolata, de Stramenopiles en de Hacrobia hebben ze verkregen door endocytobiose van Rhodophyta-plastiden.
Ze bevatten chlorofyl a + c.
Secundaire plastiden van de Euglenozoa en de Chlorarchinophyta (amoeboïde Rhizaria) zijn afkomstig van plastiden van groene algen. Eveneens door endocytobiose.
Ze bevatten chlorofyl a + b.
Sommige Dinoflagellaten bezitten sec. plastiden die afkomstig zijn van groene algen terwijl ze toch behoren tot de Alveolata.
Bij Dinoflagellaten komen zelfs tertiaire plastiden voor die afkomstig zijn van de algengroepen Bacillariophyta, Cryptophyta of Haptophyta.
Conclusie van de schrijvers van Biodiversity and Earth History: plastiden van de eukaryoten zijn afkomstig van vele onafhankelijke overgangen tussen diverse groepslijnen.
Het geslacht Paulinella is in dit opzicht wel heel bijzonder. Het behoort tot de Cercozoa (een subgroep van de supergroep Rhizaria) die cyanellen bezitten.
De chromatofoor heeft 2 membranen. Daarom is het een v.b. van onafhankelijke primaire endocytobiose.
Een cyanella is bekleed met een laagje peptideglycaan, evenals de celwand van een bacterie. Dus het is een relict van endocytobiose van een cyaanbacterie. Het bevat chlorofyl a.
Het licht wordt opgevangen door phycobillisomen. Groene planten en algen zijn dit laatste kwijt geraakt.
Het geslacht Cyanophora is hiervoor al genoemd als een levend fossiel. Het genoom wordt beschouwd als bewijs voor endocytobiose dat eenmalig gebeurde; 1 miljoen jaar geleden.

Cyanophora paradoxa
https://www.bing.com/th?id=OIP.F-hw6l-r566kPwmKBlz5MwHaE3&w=250&h=160&c=8&rs=1&qlt=90&pid=3.1&rm=2
7.3 De celstructuren binnen de Eukarya.
a Mitochondriën.
Komen voor bij alle supergroepen.
Gereduceerde mitochondriën (mitosomen en/of hydrogenosomen) komen voor bij Metazoa, Microsporidia, Chytridiomycota, Conosa, Percolozoa, Fornicata, Parabasalia, Ciliophora, Apicomplexa en Peronosporomycetes.
Dit betreft allemaal organismen die anaeroob en/of parasitair leven.
b Chloroplasten.
Primaire plastiden bij Excavata, Chlorophyta en Streptophyta met chlorofyl a en b.
Secundaire plastiden bij Euglenida en Dinophyta met chlorofyl a en b of a en c.
Bij Apicomplexa is geen chlorofyl aanwezig in de apicoplast. Ze leven parasitair.
Plastiden met een peptideglycaanlaag tussen beide membranen bij Glaucocystophyta en Paulinella met chlorofyl a.
Plastiden met een nucleomorf tussen de plastidenmembranen bij Chlorarachniophyta met chlorofyl
a en b.
Primaire plastiden bij Rhodophyta met chlorofyl a.
Secundaire plastiden afkomstig van Rhodophyta bij Chromerida en Dinophyta met chlorofyl a
of a en c.
Secundaire plastiden afkomstig van Rhodophyta waarbij de kernmembraan ribosomen bevat en zich voortzet in het endoplasmatisch reticulum (ER) en zo het chloroplast endoplasmatisch reticulum (CER) vormt. Met chlorofyl a en c.
c Celwanden.
Geen celwand: Apusozoa, Metazoa, Amoebozoa, de supergroep Excavata (uitgezonderd de Euglenida die een pellicula bezitten van eiwitten) en de supergroep Rhizaria (uitgezonderd de Foraminifera met een kalk of kiezelskelet).
Celwanden met chitine: de Holomycota.
Lorica van cellulose of chitine bij Choanomonada.
Celwanden met cellulose: de supergroep Archaeplastida.
Polysachariden: Labyrinthulida.
Cellulose en hemicellulose + chitine: Peronosporomycetes.
Siliciumoxide: Diatomeeën (Bacillariophyceae).
Celwand van silicaat of cellulose of een lorica met chitine of silicaat: goudalgen (Chrysophyceae).
Cellulose en algininezuur: bruinwieren (Phaeophyceae).
Polysachariden + meestal cellulose: Haptophyta.
Alveoli onder de celmembraan: Alveolata. Soms met ook celluloseplaten: Dinophyta.
d Reservevoedsel.
Glycogeen: de supergroepen Unikonta en de Excavata (uitgezonderd de Euglenida met paramylon en de Kinetoplastea met mannaan).
Zetmeel: Archaeplastida (uitgezonderd de Rhodophyta met
florideaan-zetmeel).
Eveneens zetmeel bij Ciliophora, Apicomplexa en Cryptophyta.
ß-glucaan: Peronosporomycetes
Chrysolaminarine: bij Bacillariophyceae (diatomeeen), Chrysophyceae (goudalgen), Phaeophyceae (bruinwieren) en Haptophyta.
Paramylon: Clorarachniophyta.
e Flagellen.
Unikont, bikont, 4 of 8 of (zeer) veel komt verspreid over alle supergroepen voor. Soms alleen de gameten of zoösporen unikont of bikont.
f Hulppigmenten.
Carotenoïde: Chlorophyta en Streptophyta.
Phycocyanine en phycoerythrine: Glaucophyta en Rhodophyta.
ß-Caroteen, neoxanrhine, diadinoxanthine en xanthophyl: Euglenida.
ß-Caroteen, fucoxanthine, violaxanthine, anthaxanthine en neoxanthine: Chrysophyceae.
ß-Carateen, fucoxanthine, diadinoxanthine en diatoxanthine: Phaeophyceae en Haptophyta.
ß-Caroteen, neoxanthine, violaxanthine, luteïne, loroxanthine-dodecanoatine: Chlorarachniophyta.
Phycoerythrine, phycocyanine en alpha-caroteen: Cryptophyta.
Fucoxanthine en peridinine: Dinophyta.
Isofucoxanthine: Chromerida.
Fucoxanthine: Bacillariophyceae (diatomeeën).
7.4 Enige conclusies uit bovenstaande.
Als ik dit geheel op me laat inwerken geeft het aanleiding tot de volgende overwegingen:
7.4.1. Het voorkomen van mitochondriën levert geen indelingscriterium op. Dat ze dankzij endocytobiose van een bacterie opgenomen zouden zijn in een eukaryote cel is voorlopig niet geloofwaardig. Tot nu toe is, voor zover mijn kennis reikt, elke opname van een 'vreemd' organisch bestanddeel in het cytoplasma onmogelijk zonder vertering m.b.v. enzymen die de organische stof bruikbaar maken voor de opbouw van het cytoplasma of voor de energiewinning. Wel komt veelvuldig voor de opname van een andere cel (bacterie, alg, schimmel of zelfs hoger organisme) als symbiont (endosymbiose).
7.4.2. Plastiden komen bij alle supergroepen voor behalve bij de Unikonta.
De chlorofylmoleculen a, b en d verschillen nauwelijks van elkaar. Bij chl. a en b is de 2e plaats (rood omlijnd) veranderd van CH3 in CHO. En bij chl. d zijn de CH3 en de CHO-groep van plaats verwisseld.
De structuurformules van chl. a, b, d, c1 en c2 zijn:
chl. a: C55 H72 O5 N4 Mg
chl. b: C55 H70 O6 N4 Mg
chl. d: C54 H70 O6 N4 Mg
chl. c1: C35 H30 O5 N4Mg
chl. c2: C35 H28 O5 N4 Mg.
Er is daarom weinig reden om het bezit van chl. a, b of d als nogal fundamenteel te hanteren. De Euglenida zouden dan goed aansluiten bij de Archaeplastida. Hoewel de hulppigmenten daar weer tegen pleiten. Ook het bezit van paramylon als reservevoedsel is een argument daartegen. Dan zou misschien aansluiting bij de Chlorachniophyta (samen met Paulinella) te overwegen zijn. De hulppigmenten passen daar ook gedeeltelijk bij. Voor beide geldt eveneens de (zeer onwaarschijnlijke) aanname dat er sprake zou zijn geweest van secundaire endocytobiose van een groene alg. Bij Paulinella zou er zelfs sprake zijn geweest van de (unieke!!) opname van een cyanobacterie.
Bij de supergroepen Chromalveolata en de Hacrobia zouden de plastiden afkomstig zijn van de Rhodophyta dankzij secundaire en tertiaire endocytobiose.
Bij Stramenopiles en Hacrobia is er sprake van CER: chromoplast-endoplasmatisch-reticulum. Is dit een erg fundamenteel-specifieke celconstructie?
Veel nader onderzoek zou vereist zijn om eventueel duidelijk te maken waarom er 2, 3 of 4 membranen per plastide belangrijk zijn voor het functioneren daarvan.
Lettend op bovenstaande structuurformules lijkt het van belang dat chl. c (c1 en c2?) voorkomen bij de Chromalveolate en de Hacrobia. Bij beide supergroepen en de Rhizaria komen ook een rijke variatie (en deels overeenkomstige) hulppigmenten voor. Ook dat vraagt nader onderzoek als het gaat over de ecologische plaats binnen het leefgebied.
7.4.3. De bouw van celwanden vertoont een opmerkelijke variatie binnen de supergroepen. Vele groepen van de Unikonta bezitten chitine. Vandaar dat binnen deze supergroep de "schimmels" (Holomycota) worden verenigd met de Metazoa. Omdat de Arthropoda (geleedpotigen) ook chitine hebben. Is dit doorslaggevend ?
De Apusozoa missen chitine maar deze groep wordt door de geleerden in heel verschillende supergroepen geplaatst. Ze hebben een theca van .....? De Choanomonada hebben een lorica van cellulose en silicium.
De Amoebozoa zijn naakt of zijn bekleed met organisch materiaal of met mineralen. Amoeboïde vormen en slijmzwammen komen voor in diverse (super)groepen en passen m.i. alleen binnen de Unikonta vanwege het bezit van reservevoedsel glycogeen.
7.4.4. Glycogeen als reservevoedsel komt in vele uiteenlopende groepen voor van amoebe-achtigen en schimmels t/m alle Metazoa (o.a. de mens). Bovendien nog bij de supergroep Excavata. De laatste is toch al een merkwaardig fenomeen: Ze leven merendeels anaeroob en hebben geen plastiden of mitochondriën. Sommige leven amoeboïd of als slijmzwam. Ze hebben geen, 2 of vele flagellen.
Ze leven vaak als commensaal of parasitair. Een achterwaarts gerichte flagel voert water met prooi naar de cytosoom. Maar of dit nu een scheidend criterium is?
Zeer opvallend: de Euglenida vinden hierbinnen een plek. Wel, naast zetmeel, met een ander reservevoedsel: paramylon (zie boven).
Zetmeel wordt ook opgeslagen in de cellen van de Alveolata. Dit is ook het geval bij de Cryptophyta. Maar bij de Bacillariophyceae (de diatomeeën), de Chrysophyceae (de goudalgen) en de Phaeophyceae (de bruinwieren) bestaat het uit chrysolaminarine (zie boven). Ook ß-glucaan komt voor.
Al met al lijkt me het type voorkomend reservevoedsel (allemaal polysachariden) niet kenmerkend voor de indeling in (super)groepen.
7.4.5 Het indelingscriterium 'aantal flagellen' is niet erg bruikbaar. Alle mogelijke variaties komen binnen alle super- en subgroepen voor. Bovendien is het zeer de vraag of b.v. de duizenden ciliën bij de Ciliophora (Ciliata), die alle perfect met elkaar samenwerken, wel vergelijkbaar zijn met de flagellen bij andere supergroepen. Daarvoor is er meer kennis nodig.
7.4.6 Het aantal mogelijke hulppigmenten is zeer divers. Bij alle groepen groenwieren, roodwieren, bruinwieren, dinofyten, diatomeeën en andere groepen 1-celligen zou je moeten bestuderen welke delen van het licht precies worden geabsorbeerd i.v.m. de ecologische positie van die organismen. Als resultaat daarvan zou er wellicht een andere indeling van al deze groepen waarschijnlijker blijken te zijn dan nu wordt aangenomen.
Hoofdstuk 8 Enkele samenvattende conclusies.
1. Zonder de hypothese van het proces endocytobiose valt het hele cladistische systeem in elkaar.
Afzonderlijke processen van endocytobiose zouden door experimenten aangetoond behoren te worden. Tot nu toe heb ik daar niets over gelezen. Er bestaat een vakgroep experimentele evolutie. Wellicht bieden resultaten binnen die groep nog eens boeiende resultaten hieromtrent.
De endosymbiose-hypothese veronderstelt nu dat er kernvorming plaats vond door het samengaan van een bacterie met een pro-eukaryotische cel (een Archea). De bacterie verloor b.v. z'n celwand (Hoe is dat mogelijk?) en slokte een Archea op. Daarna verloor de Archea z'n celwand en plasmamembraan (hoe kan dat?). Tegelijkertijd vormde de gastheerbacterie instulpingen in de celmembraan. Eventueel werd het gastheer-genoom overgeplaatst in de Archea en daarin werd een kern en endoplasmatisch reticulum (ER) gevormd. Aldus het gefingeerde verhaal in Microbiology door Presscott, Harley en Klein.
We leren hieruit: biologische fantasie staat voor niets!!
De nieuwste favoriet voor deze "geschiedenis" is Prochloron. Deze cellen bevatten chlorofyl a en b. Een veronderstelde voorouder van Prochlorophyten en een cyanobacterie zouden samen een chloroplast gevormd kunnen hebben.
De aldus ontstane primitieve eukaryotische cel zou samen met bacteria een mitochondrium gevormd kunnen hebben. En zo ontstonden de eerste pro-chlorophyten (primitieve groene algen).
Zowel bladgroenkorrels als mitochondriën vertonen een ingewikkelde membranenstructuur waarin zeer gecompliceerde stofwisselingsprocessen verlopen. Opname van (cyano)bacteriën geeft hiervoor geen verklaring. Die onverklaarbare hypothese verschuift trouwens alleen maar het probleem. En dan rest nog de verklaring voor het ontstaan van een perfect georganiseerd Endoplasmatisch Reticulum in absolute samenhang met de kern en haar chromosomen.
2. De diverse cladistische vertakkingsschema's met de onderlinge afstanden tussen de takken en de variaties in de lengtes van die takken is onbevredigend onderbouwd. Berekeningen gebaseerd op de frequenties van nucleïnezuren en de onderlinge afstanden daartussen in het chromosoom lijken onbetrouwbaar en vaak onderling tegenstrijdig. De veronderstelde samenhang hiervan met vele stofwisselingsprocessen biedt geen oplossing voor bewijzen van ontstaan van groepen en onderlinge verwantschappen. Zelfs de sterkste computers vermogen geen uitsluitsel daaromtrent te geven!
3. Het hele terrein van 1-cellige "algen" met of zonder chloroplasten, met of zonder skelet, met of zonder zetmeel of cellulose, met of zonder flagellen of trilharen vormen een bijna onafzienbaar geheel van veronderstelde monofyletische, polyfyletische of parafyletische groepsindeling. Afgezien van redeneringen rond endocytobiose zijn nergens overtuigende evolutielijnen tussen de hoofdgroepen of subgroepen te trekken. Alle mooie cladistische lijnen (ook op het internet, Wikipedia) vragen voorafgaand geloof en een goed vertrouwen op de "wetenschap" van het systeem.
4. Als we van waterplanten, evolutionair gezien, overgaan naar landplanten is er sprake van wensdenken. De onder de Streptophyta genoemde zgn. voorbereidingen wijzen daar op!
Naast cuticula, huidmondjes en rhizoiden moeten diverse blad-, stengel- en wortelsystemen ontstaan.
Ontwikkelingslijnen, met en zonder blijvend cambium, zijn gemakkelijk te trekken. Maar hoe je van (lever)mossen varens maakt en van die laatste weer paardenstaarten en zaadplanten blijft volledig onopgelost. De noodzakelijke veranderingen zijn zo fundamenteel samenhangend dat geen enkele geleerde tot nu toe een goed onderbouwd verhaal daarover heeft weten te maken. Vooral hoe dubbele bevruchting en de vorming van reservevoedsel in zaadlobben en de bouw van zaden en vruchten hebben kunnen ontstaan behoort nog altijd tot natuurraadsels of -wonderen!
5. De indeling in 6 supergroepen is erg onbevredigend. Als we letten op de bouw en structuur van cellen, weefsels en organen is het hoogst merkwaardig om de Holomyceta in de supergroep van Unikonta te plaatsen. Samen met alle "diergroepen" t/m de Primaten.
Even merkwaardig is de plaats van Phaeophyceae, Chrysophyceae en Bacillariophyceae samen met Ciliophora (o.a. pantoffeldiertje) en Dinophyta in de supergroep Chromalveolata.
Wellicht zou het logisch zijn om alle organismen zonder weefselstructuren binnen 1 supergroep te plaatsen. Misschien zou verdere studie van celstructuren, celwanden, reservevoedsel, en stofwisselingsprocessen kunnen leiden tot een verdere indeling van een dergelijke groep.
De Porifera (sponzen) zouden daarbij een plek op de valreep kunnen innemen.
Amoeboïde cellen en slijmcellen zouden samen in 1 super(sub)groep kunnen worden geplaatst evenals cellen met versterkte celmembranen door cellulose, kalk- of kiezelskeletten.
Het bezit van chloroplasten zou kunnen fungeren als een secundair criterium voor de indeling; evenals het bezit van chlorofyl c1 of c2.
6. Letten we op mogelijke tijden van ontstaan van de grote "plantenafdelingen" dan blijkt het volgende:
Het ontstaan van alle bacteriën en eukaryoten vanuit de levenloze materie is principieel onbewijsbaar.
Zowel binnen de Archaeoplastida (de "planten") als de Holozoa ( de "dieren") worden gigantische evolutionaire sprongen gemaakt zoals b.v. tussen "algen en mossen", tussen "mossen en varens", tussen "varens en zaadplanten", tussen "naaktzadigen en bedektzadigen", tussen 1- en 2-zaadlobbhigen, tussen "sponzen, holtedieren en ribkwallen", tussen "trilobieten, krabben, spinnen, insecten en schorpioenen", tussen "platwormen, raderdiertjes, armpotigen, ringwormen en weekdieren", tussen "graptolieten en stekelhuidigen", enz. enz. Enig fossiel bewijs is daarbij meestal niet gevonden.
7. Alle grote groepen "water- en landplanten" bestaan voor het Krijt.
Als we er van uitgaan dat. tegen het einde van het Krijt de grootste van de 5 extinctieperiodes plaats greep en alle .grote groepen organismen gedurende de periode Precambrium t/m Krijt bestonden vanaf het begin van de schepping, betekent dit dat alle grote afdelingen van het plantenrijk t/m de tweezaadlobbige zaadplanten gedurende alle aardperioden aanwezig waren.
Gedurende het Trias wordt verondersteld dat Pangea bestond. Met de onderdelen Gondwana en Siberia. Er heerste toen een gelijkmatig warm klimaat en grote ondiepe zeegebieden langs de kusten. Alle groepen hoogontwikkelde planten (Gymnospermen, zaadvarens en zaadplanten) en dieren (tweekleppigen, geleedpotigen, inktvissen, (kraak)beenvissen, amfibieën, reptielen (Sauriërs) en zoogdieren) worden in die periode als fossiel gevonden. Terwijl aan het einde van het Perm enorme vulkaanuitbarstingen (en andere processen) zorgden voor het uitsterven van vele zaadplanten en (zee)dieren. Tijdens de Jura blijft het klimaat tamelijk rustig. Alleen iets koeler. Bomen krijgen jaarringen! Verondersteld wordt dat er nu grote zoutlagen en aardolie en aardgas zijn ontstaan.
Dat laatste vooral dankzij enorme productie van plankton en nekton in de ondiepe zeeën.
De continenten beginnen uiteen te drijven. Als dat proces vrij snel verliep ging het gepaard met orogenese. In de geologie worden 5 perioden van orogenese genoemd. Ik neem aan dat orogenese-processen samengaan met grote extincties en vrij snel verliepen (zie daarvoor de kaartjes in Hoofdstuk 4.2).
Tijdens het Krijt valt Pangea verder uiteen.
1 Noordelijke Atlantische oceaan 2 Zuidelijke Atlantische oceaan
3 Thethys oceaan.
geel gestippeld: ondiepe kustzeeen.
Als de continentverschuivingen vrij snel hebben plaats gevonden zijn er in die periode ook vele, misschien lagere gebergten ontstaan. Dat zal gepaard zijn gegaan met aardbevingen, vulkaanuitbarstingen en mogelijk vloedgolven. Tijdens dergelijk gebeurtenissen kunnen gigantische zoutlagen, aardolie en aardgas zijn gevormd. Steenkool kan daarvoor en daarna zijn ontstaan.
Ontstaan van onvoorstelbaar grote hoeveelheden (biljoenen of triljoenen tonnen) steenzout, aardolie, aardgas en steenkool kan onmogelijk plaats gegrepen hebben tijdens kleine stapjes van enkele cm's sedimentatie per jaar. Het lijkt me evenmin een reële veronderstelling dat dit allemaal tijdens, kort voor of kort na de zonvloed zal zijn gebeurd.
Bedektzadigen (waaronder grassen) zijn gefossiliseerd vanaf de periode Krijt. Evenals vogels (zij vervangen de Pterosauriers) en zoogdieren (van knaagdieren tot ......). 70% van alle bloemplantensoorten groeien alom dankzij de bestuiving door insecten! Ook termieten en mieren zijn aanwezig.
1 Gondwana 2 Laurentia 3 Siberia 4 Baltica
De verdeling van het land tijdens het Cambrium zoals weergegeven lijkt me erg hypothetisch. Ze zal ongetwijfeld ontstaan zijn n.a.v. fossielen die aan die geologische periode worden toegeschreven en daarvoor. Ik denk dat het veel beter is om de gevonden fossielen in te delen op basis van ecologische leefgebieden. De organismen tijdens precambrium t/m/ Devoon waren de Ediacara-fauna (veelal zonder harde delen; bedolven onder vulkanische as), de eerste schelpdieren Graptolieten en Arthropoden, zeeschorpioenen (Eurypteriden), zeeanemonen, Brachiopoden, wormen, koralen, zeelelies, zeesterren, schelpdieren, slakken, inktvissen, kaakloze vissen (Agnatha), Placodermi, haaien, longvissen, Sarcopterygiers, Tiktaalik en Conodonten. Op het land leefden Cooksonia, Rhynia, Asteroxylon (echte vaatplanten) Archaeopteris (bomen tot 18m. hoog), korstmossen, chlorophyten, Ichtyostega (een zwemmer met kieuwen en zwempoten), spinachtigen en duizendpotigen.
Het leven speelde zich af in ondiep water (tot 50 m.) langs de kusten overal ter wereld. En aansluitend daarbij op kusten met dichte plantengroei van sporenplanten met generatiewisseling van sporofyt en gametofyt (Psilofyten, Lycofyten, Equisetae).
Tijdens de periode van het Perm begint de verdeling van land en water al veel te lijken op die van het
Het Paleogeen.
1 Antarctica 2 Zuid-Amerika 3 Noord-Amerika
4 Groenland 5 "Europa" 6 Siberië 7 China
8 India 9 Afrika 10 Australië.
De aardbol zou na de schepping grote overeenkomst kunnen vertonen met die van Pangea.
Al de gegevens en fossiele vondsten van de ecologische gebieden benoemd als precambrium, ediacara, cambrium, ordovicium, siluur en devoon kunnen met elkaar samenvallen. Ingrijpende gebeurtenissen op land- en zeegebieden zoals orogeneseprocessen en als gevolg daarvan bedekking met enorme lagen sedimenten van opeengehoopte lagen plantaardig en dierlijk materiaal kunnen vanaf het paradijsleven hebben plaats gevonden. Na het Devoon begint Pangea langzaam uiteen te vallen.
Alleen dergelijke gebeurtenissen kunnen het ontstaan van de vele steenkoollagen, de schier eindeloze voorraden aardolie en aardgas en de enorme zoutlagen in vele wereldgebieden verklaren.
Fossilisatie van grote, snel zwemmende waterdieren en van snel vliegende insecten en vogels is totaal onbegrijpelijk zonder de aanname van snel verlopende ingrijpende uitbarstingen van gewelddadige gebeurtenissen. Trouwens ook het bedolven of ingevroren raken van middelgrote en zelfs kolossale landdieren (buideldieren, dinosauriers, zoogdieren en reptielen) is niet te begrijpen vanuit langzaam verlopende processen.
Ondanks deze veronderstellingen blijven er vele onopgeloste vragen bestaan.
Waardoor zijn er vindplaatsen waarin geen fossielen van Benettitales, Cycadeeën, Coniferen en Angiospermen voorkomen?
Eenzelfde probleem betreft het niet voorkomen van grote groepen gewervelde dieren zoals Actinopterygiers, Amfibieën, Reptielen, Vogels en zoogdieren.
Nieuwe ontdekkingen en onderzoekingen zijn hiervoor noodzakelijk. Daarbij dient rekening te worden gehouden met eigenschappen van die groepen organismen en de omstandigheden waaronder fossilisatie plaats greep.
Vooral van belang is rekening houden met het voorkomen van vele groepen landdieren en landplanten op de enorm uitgestrekte niet onderzochte landdelen van Pangea.
Reageren? tnbuist@gmail.com









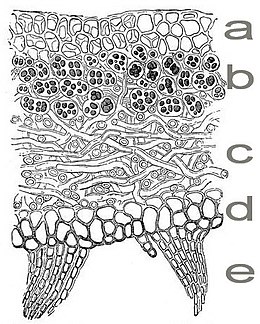


 Sporangia aan de onderzijde van een varenblad. Ze zijn vaak bedekt met een vlies om de sporangia te beschermen. De sporangia hebben een annulus om ze open te scheuren bij droogte om de sporen vrij te laten.
Sporangia aan de onderzijde van een varenblad. Ze zijn vaak bedekt met een vlies om de sporangia te beschermen. De sporangia hebben een annulus om ze open te scheuren bij droogte om de sporen vrij te laten.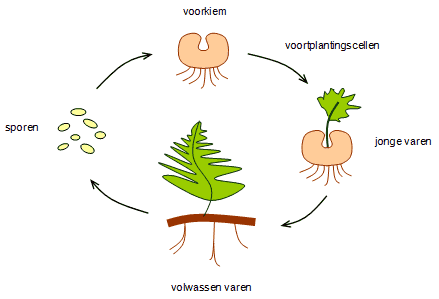
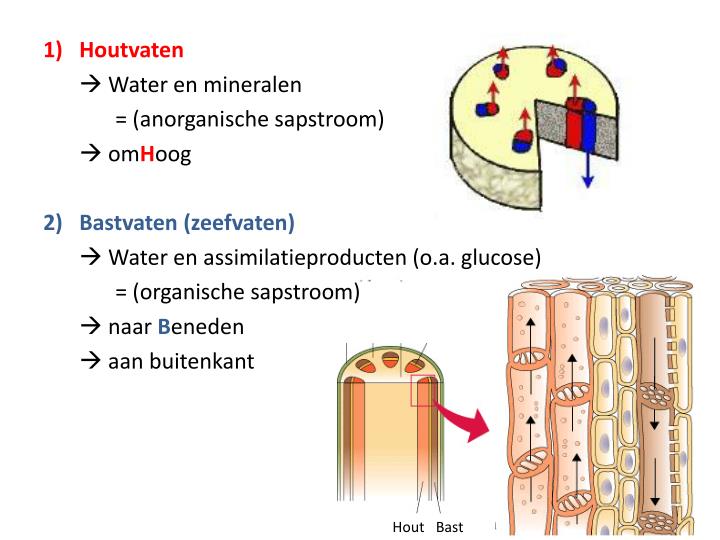















 Anomalocaris. bron Wikipedia.
Anomalocaris. bron Wikipedia.
 Een Conodont. bron Wikipedia.
Een Conodont. bron Wikipedia. Pterygotus, een Eurypteride. Jagend op vissen. bron Wikipedia.
Pterygotus, een Eurypteride. Jagend op vissen. bron Wikipedia.

 Eustenopteron. bron Wikipedia.
Eustenopteron. bron Wikipedia.
 Acanthostega. bron Wikipedia.
Acanthostega. bron Wikipedia.





















































 Watongia. Bron Wikipedia.
Watongia. Bron Wikipedia.















































 bron Wikipedia.
bron Wikipedia.




 Chytridiomycota.
Chytridiomycota.



 Glaucocystophyta.
Glaucocystophyta.


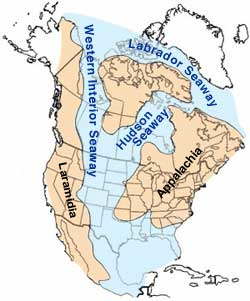{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





 Apicoplexa
Apicoplexa Chrysophyceae.
Chrysophyceae.






Reacties
Een reactie posten